Intelligent Design
Theory ľ An Overview
Peter S. Williams
(MA, MPhil)
Intelligent Design (ID) is a scientific research program embraced by a relatively small but
growing group of scientists and other academics (called Ĺdesign theoristsĺ) who
argue that: Ĺintelligent agency, as an aspect of scientific theory making, has
more explanatory power in accounting for the specified, and sometimes
irreducible complexity of some physical systems, including biological entities,
and/or the existence of the universe as a whole, than the blind forces of. . .
matter.ĺ[1]
That is, intelligent design is a better explanation for entities exhibiting complex
specified information (CSI)[2]
than are appeals to the inherent capacities of nature (i.e. chance and/or
physical necessity). ID suggests that the world contains objects that exhaust
the explanatory resources of undirected natural causes, and can only be
adequately explained by recourse to intelligent causation. For example,
according to biochemist Michael J. Behe:
itĺs been the very progress of science itself that has made
intelligent design plausible. Fifty years ago much less was known about the
cell, and it was much easier then to think that Darwinian evolution was true.
But with the discovery of more and more complexity at the foundation of life,
the idea of intelligent design has gained strength. That trend is continuing.
As science pushes on, the complexity of the cell is not getting less; on the
contrary, it is getting much greater.[3]
Design theorists have claimed that intelligent design can be inferred from:
- The
fine-tuning of the laws of nature[4]
- The
fine-tuning of our cosmic habitat (e.g. the structure of our solar system)[5]
- The origin of
life (e.g. DNA)[6]
- Irreducibly
Complex bio-molecular machines (e.g. the bacterial flagellum)[7]
- The Cambrian
Explosion[8]
Philosopher Francis J. Beckwith reports that: ĹID proponents have developed highly
sophisticated arguments, have had their work published by prestigious presses
and in academic journals, have aired their views among critics in the corridors
of major universities and institutions, and have been recognized by leading
periodicals, both academic and non-academic.ĺ[9]
 |
Perhaps the best single volume introduction to intelligent design theory is Science and Evidence for Design in the Universe (Ignatius Press, 2000), a collection of densely argued papers by leading design theorists biochemist Michael J. Behe, mathematician and philosopher William A. Dembski & philosopher of science Stephen C. Meyer.
[Click on cover picture to purchase. Cover pictureĺs from www.amazon.co.uk] |
ID is Not ĹCreation Scienceĺ[10]
ĹIntelligent design theory. . . says nothing about the Bible.ĺ[11]
Oxford zoologist Richard Dawkins lumps ID together with Ĺcreation
scienceĺ, calling ID a Ĺeuphemism for creationists.ĺ[12] Leonard Krishtalka notoriously
slighted ID as Ĺcreationism in a cheap tuxedo.ĺ[13] This rhetorical slight of hand
is a consistent feature of Darwinian apologetics: Ĺanyone critical of Darwinian
evolution risks being stereotyped as a strict believer in biblical creation.ĺ[14] Paleontologist Stephen Jay
Gould asserted that the only dissenters from macroevolution are Ĺprotestant
fundamentalists who believe that every word of the Bible must be literally
true.ĺ[15]
In reality: Ĺsome of the strongest critics of Darwinĺs theory are scientists
who happen to be non-fundamentalist Protestants, Catholics, or Jews (as well as
agnostics).ĺ[16]
As John Angus Campbell says:
The ID movement, comprising as it does academics, scientists,
philosophers, humanist educators, and interested laypeople, is certainly not
the same, except for purposes of histrionic exaggeration, as the young-earth,
six-literal-days Ĺcreation scienceĺ of the past. . . In the ambiance of IDĺs
Ĺbroad tentĺ. . . one will find persons of many philosophic perspectives and
metaphysical commitments.[17]
Historian of science Ronald L. Numbers observes that proponents of
evolution tend to conflate ID and creationism: ĹThey see intelligent design as
little more than gussied up creationism, despite the significant differences.ĺ[18] However, as Dr. Benjamin Wiker
observes: Ĺintelligent design theorists who are Christians are very careful to
distinguish themselves from creationists.ĺ[19]
This is not because design theorists necessarily accept Darwinistĺs low
opinion of Ĺcreationismĺ. Indeed, some design theorists are also
Ĺcreationistsĺ; just as some evolutionists are also atheists. However,
ID is not Ĺcreationismĺ (anymore than evolution, as opposed to naturalistic
evolution or Darwinism, is inherently atheistic). The story told by ID:
veers away from the usual theistic evolutionary story (Ĺbased
on the evidence, theistic scientists are now concluding that God worked through
evolutionĺ) and from the classic creation science tale (Ĺscientists are
recognizing that Genesis is literally true after allĺ).[20]
Hence the need to draw clear lines of demarcation and to avoid making criticisms of the former
that, whether sound or unsound, could only apply to the latter.
Design theorist, philosopher and mathematician William A. Dembski is
at pains to stress that ID is not Ĺcreationismĺ as popularly understood:
the design theoristsĺ critique of Darwinism in no way hinges on the
Genesis account of creation. . . The design theoristsĺ beef is not with
evolutionary change per se, but with the claim by Darwinists that all
such change is driven by purely naturalistic processes. . . the design
theoristsĺ critique of Darwinism begins with Darwinismĺs failure as an
empirically adequate scientific theory, and not with its supposed
incompatibility with some system of religious belief.[21]
Design theorist Professor Phillip E. Johnson stresses: ĹI am not a
defender of creation-science. . . The essential point of creation has nothing
to do with the timing or the mechanism the Creator chose to employ, but with
the element of design or purpose.ĺ[22]
Johnsonĺs 1989 ĹPosition Paper on Darwinismĺ explains: ĹThe truly fundamental
question is whether the natural world is the product of a pre-existing
intelligence and whether we exist for a purpose that we did not invent
ourselves.ĺ[23]
Johnson points out that:
Creationists are not necessarily Genesis literalists or believers in
a young earth, nor do they necessarily reject Ĺevolutionĺ in all senses of that
highly manipulable term. A creationist is simply a person who believes. . .
that the living world is the product of an intelligent and purposeful Creator
rather than merely a combination of [unintended] chance events and impersonal
natural laws.[24]
In Johnsonĺs sense of the word, theistic evolutionists (who accept evolution as Godĺs method
of creation via secondary causes) are creationists, and a creationist neednĺt
even be a theist. They certainly donĺt need to accept any of the various
interpretations of the Genesis creation story.[25] They simply need to accept the
metaphysical hypothesis of design.
 |
In the influential bestseller Darwin on Trial (IVP, 1993), the Jefferson Peyser Emeritus Professor of law at the University of California at Berkley, Phillip E. Johnson, argues that the theory of evolution is based on faith in naturalism rather than upon sufficient scientific evidence. |
Design theorists additionally accept design as a scientific inference to the best
explanation: Ĺdesign theorists do not defend their position by appealing to
esoteric knowledge, special revelation, or religious authority. They make
philosophical and scientific arguments whose merits should be assessed by their
soundness. . .ĺ[26]
Dr. Thomas Woodward, author of Doubts about Darwin: A History of Intelligent
Design, repudiates the claim that ID is motivated by religious premises:
hearing how key Design advocates came to their current view,á it
became clear that their entry into the movement stemmed from intellectual or
scientific ľ not religious ľ reasons. . . Several of the founders frequently
relate a vivid tale of how they previously had assumed the validity of
Darwinian scenarios and were later shocked to discover major weaknesses in the
case for Darwinism.á Typically this intellectual epiphany leads to further
reading and research, which cements the new radical doubt about the theoryĺs
plausibility.[27]
 |
Professor Thomas Woodwoodĺs Doubts about Darwin: A History of Intelligent Design (Baker, 2003) is an excellent history of and introduction to the intelligent design movement, with a foreword by Phillip E. Johnson. |
ID suggests that the religiously unencumbered concept of intelligent design is a better and more
fruitful explanation of certain biological and physical evidence than the
hypothesis that natural regularities and chance (whether or not these causes
are themselves created) are responsible.
ID is Not Natural Theology[28]
ĹID is a research program whose inferences support, and are consistent with, some
belief in a higher intelligence or deity; it is not a creed that contains
belief in a specific deity as one of its tenets.ĺ ľ Francis J. Beckwith[29]
While a design paradigm has historically dominated cosmology and biology (from Anaxagorus,
Socrates, Plato and Aristotle through to Newton and Paley), design arguments
have also been a part of natural theology, the philosophical project of
providing evidence for Godĺs existence. However, design arguments can only
provide part of the evidence for God.[30]
The ID movement recognizes this fact and insists upon distinguishing between
arguing for intelligent design and arguing for divine design (a
fact that critics of ID often obfuscate). Behe writes:
my argument is limited to design itself; I strongly emphasize that
it is not an argument for the existence of a benevolent God, as Paleyĺs was. I
hasten to add that I myself do believe in a benevolent God, and I recognize
that philosophy and theology may be able to extend the argument. But a
scientific argument for design in biology does not reach that far. Thus while I
argue for design, the question of the identity of the designer is left open. .
.á as regards the identity of the designer, modern ID theory happily echoes
Isaac Newtonĺs phrase, hypothesis non fingo.[31]
ID argues that there are natural entities that cannot reasonably be explained by chance and/or
physical necessity, and only then infers, on the basis of experience, that the
best explanation of these features of reality is intelligent causation (ID is
not an Ĺargument from ignoranceĺ). Whether the intelligence in question is God
is yet a further question: Ĺdetecting design. . . does not implicate any
particular intelligence.ĺ[32]
Failure to appreciate the distinction between intelligent design and
divine design has contributed to the scientific establishment throwing out the
design paradigm as essentially tied to belief in God when it was not. After
all, one could accept intelligent design and attribute it to the activity of
angels, demons, Platoĺs finite god (the Demiurge), the gods of Egyptian, Greek
or Norse polytheism, or to aliens, rather than to God (hence it is
logically possible for an atheist to support ID[33]).
The plain truth
is that ĹIntelligent design is not a form of natural theologyĺ[34]; and while every design
argument for God is an argument for intelligent design, not every argument for
intelligent design need be viewed as an argument for God - at least, not
without considerations from outside ID as a scientific theory being brought to
bear: Ĺintelligent design theory by itself makes no claims about the nature of
the designer, and scientists currently working within an intelligent design
framework include Protestants, Catholics, Jews, agnostics, and others.ĺ[35] As Benjamin Wiker suggests:
Ĺfrom scientific evidence open to all, we can infer that nature has an
intelligent designer. Further, we can extend these arguments philosophically,
demonstrating that the intelligent designer is God.ĺ[36]
 |
A good example of natural theology building upon intelligent design arguments is Journalist Lee Strobelĺs The Case for a Creator (Zondervan, 2004), in which that author interviews a number of scholars associated with the ID movement (including William Lane Craig, Stephen C. Meyer, J.P. Moreland Guillermo Gonzalez, Jay Richards & Jonathan Wells) as he builds a cumulative case for the existence of a Creator. |
ID obviously has much to contribute to natural theology, in that divine design is
a plausible and independently supported candidate for the source of
scientifically detectable intelligent design. Theologically speaking, it is
interesting to note, given IDĺs emphasis on the need to explain the origin of
information that the Bible portrays God as creating through his Word or ĹLogosĺ
(Gen 1:3, John 1:1-3): ĹOf course God did not physically speak and produce
sound wavesĺ, writes theologian Terence L. Nichols, ĹBut the point is that the
word conveys information. Gods act of creating. . . involves the input of
specifying information. . .ĺ[37] Likewise, natural theology has much to contribute to ID. The two projects are
mutually re-enforcing. On the one hand, ID furnishes natural theology with the
basis for powerful new versions of the design argument. On the other hand,
anyone who is already impressed by natural theology should be kindly
predisposed towards ID.
Part I: ID Theory -
Admitting the Possibilities & Following the Evidence
 In Laws (Book X) the Greek philosopher Plato observed that:
Ĺall things do become, have become and will become, some by nature, some by art
[intelligent design], and some by chance.ĺ[38]
ID constitutes a scientific paradigm based upon acknowledging all of the
explanatory possibilities listed by Plato and the existence of reliable
criteria legitimating evidentially motivated inferences to the explanatory
superiority of intelligent design (Ĺartĺ) in certain well-defined circumstances,
thereby freeing science to follow the evidence wherever it leads.
In Laws (Book X) the Greek philosopher Plato observed that:
Ĺall things do become, have become and will become, some by nature, some by art
[intelligent design], and some by chance.ĺ[38]
ID constitutes a scientific paradigm based upon acknowledging all of the
explanatory possibilities listed by Plato and the existence of reliable
criteria legitimating evidentially motivated inferences to the explanatory
superiority of intelligent design (Ĺartĺ) in certain well-defined circumstances,
thereby freeing science to follow the evidence wherever it leads.
(Plato, from The School of Athens by Raphael[39])
The Ĺopenĺ philosophy of science[40]
adopted by ID is in stark contrast to the approach of those who work within the
explanatory confines of metaphysical or even Ĺmethodologicalĺ naturalism.
According to Richard Dawkins: ĹThe kind of explanation we come up with must not
contradict the laws of physics. Indeed it will make use of the laws of
physics, and nothing more than the laws of physics.ĺ[41] Richard Lewontin openly
admits:
It is not that the methods. . . of science somehow compel us to
accept a material explanation of the. . . world, but, on the contrary, that we
are forced by our. . . adherence to material causes to create. . . a set of
concepts that produce material explanations, no matter how counterintuitive, no
matter how mystifying. . .[42]
But as William A. Dembski notes, here: Ĺwe are dealing with a naturalistic metaphysic that
shapes and controls what theories of biological origins are permitted on the
playing field in advance of any discussion or weighing of evidence.ĺ[43] And as philosopher of science
Del Ratzsch observes:
The scientific attitude has usually been characterised as a
commitment to following the evidence wherever it leads. That does not look like
promising ammunition for someone pushing an official policy of refusing to
allow science to follow evidence to. . . design no matter what the evidence
turns out to be. . . it commits science to either having to deliberately ignore
major (possibly even observable) features of the material realm or
having to refrain from even considering the obvious and only workable
explanation, should it turn out that those features clearly resulted from
[intelligent] activity. . . any imposed policy of naturalism in science has the
potential not only of eroding any self-correcting capability of science but of
preventing science from reaching certain truths. Any imposed policy of
methodological naturalism will have precisely the same potential consequences.[44]
If philosophical naturalism is true, then a policy of methodological naturalism cannot possibly
subvert the truth seeking intent of science; but perhaps philosophical
naturalism is not true, and perhaps science should operate without making any
assumptions that might force it to ignore reality.
 |
Science & Itĺs Limits: The Natural Sciences in Christian Perspective by Professor Del Ratzsch (Apollos, 2000) is a readable and comprehensive introduction to the philosophy of science. |
At the very least, the rule of Ĺmethodological naturalismĺ should be stated so as to make a
distinction between recognising intelligent design and supernatural design: Ĺin scienceĺ, says Larry Witham, Ĺthe question is not between finding
natural causes or supernatural causes, but between natural and intelligent
ones.ĺ[45] (It should be noted that this distinction ľ which I have called the distinction
between Ĺhard-lineĺ and Ĺsoftĺ methodological naturalism ľ sidesteps the
question of whether intelligence is by its very nature a supernatural reality.[46] Hence, one could think of this
distinction as a methodological distinction allowing ID to stake out the
broadest possible Ĺtentĺ by leaving the ontological nature of intelligence
outside of its core commitments.)
Besides: Ĺrecent studies in the philosophy of science have
confirmed. . . that philosophically neutral criteria that can define science
narrowly enough to disqualify theories of creation or design without also
disqualifying Darwinism and/or other materialistic evolutionary theories on
identical grounds do not exist.ĺ[47]
And as philosopher of science J.P. Moreland points out:
some branches of science, including SETI, archaeology, forensic science,
psychology and sociology, use personal agency and various internal states of
agents (desires, willings, intentions, awareness, thought, beliefs) as part of
their description of the causal entities, processes, events or actions cited as
explanations for certain phenomena. . . Thus there is nothing non-scientific
about appealing to personal agency and the like in a scientific explanation[48]
Indeed, many sciences depend upon detecting intelligence: Ĺnotably forensic science,
artificial intelligence. . . cryptography, archaeology and the Search for
Extraterrestrial Intelligence [SETI].ĺ[49]
ID simply suggests: Ĺthat we extend these insights, which have proved so
fruitful in other fields, to the world of the natural sciences.ĺ[50]
Darwinist Michael Ruse acknowledges that: ĹIt would indeed be very
odd were I and others to simply characterize ôscienceö as something which, by
definition, is based on (methodological) naturalistic philosophy and hence
excludes [intelligent design].ĺ[51]
In a speech given at the annual meeting of the American Association for the
Advancement of Science in 1993, Ruse conceded that: Ĺphilosophically one should
be sensitive to what I think history shows, namely, that evolution. . .
involves making certain a priori or metaphysical assumptions, which at some
level cannot be proven empirically.ĺ[52]
Little wonder, then, that: ĹA central claim of the ID movement is that if
science education is to be other than state-sponsored propaganda, a clear and
principled distinction must be drawn between empirical science and the
materialist philosophy that drives contemporary Darwinian theories of
biological origins.ĺ[53]
According to design theorists, developments in information theory
mean that there now exist Ĺwell-defined methods that, on the basis of
observational features of the world, are capable of reliably distinguishing
intelligent causes from undirected natural causes.ĺ[54] Hence William A. Dembski
defines intelligent design as: Ĺthe science that studies how to detect
intelligence.ĺ[55]
Thus, according to Dembski, ID is properly formulated as a theory of
information:
intelligent design is therefore not the study of intelligent causes per
se but of informational pathways induced by intelligent causes. As a result
intelligent design presupposes neither a creator nor miracles. . . it detects
intelligence without speculating about the nature of the intelligence. . .[56]
As Geoscientist Marcus R. Ross explained in a presentation before the Geological Society of
America: ĹID is classified as a philosophically minimalistic position,
asserting that real design exists in nature and is empirically detectable by
the methods of science.ĺ[57]
It is this minimal metaphysical baggage that keeps ID firmly within the realm
of science.
Circular Arguments
Scientists who allow a commitment to naturalistic explanations to predetermine their
explanatory options when faced with empirical evidence of intelligent design
are, to use an example from physicist Robert Kaita, like researchers stubbornly
committed to finding a naturalistic explanation for Ĺcrop-circlesĺ:
 mysterious patterns were found in
wheat fields taking the form of large, distinct geometric patterns. . .
speculation ran from intelligent causes, such as ingenious pranksters, or the
perennial favorite, extraterrestrial beings, to natural phenomena. Finally a
couple of men admitted responsibility and revealed that their equipment
consisted of very large versions of the stylus and string that people have been
using to make geometric figures since antiquity. . . In spite of this unequivocal
evidence for design, some [people] persisted in suggesting highly improbable
natural causes.[58]
mysterious patterns were found in
wheat fields taking the form of large, distinct geometric patterns. . .
speculation ran from intelligent causes, such as ingenious pranksters, or the
perennial favorite, extraterrestrial beings, to natural phenomena. Finally a
couple of men admitted responsibility and revealed that their equipment
consisted of very large versions of the stylus and string that people have been
using to make geometric figures since antiquity. . . In spite of this unequivocal
evidence for design, some [people] persisted in suggesting highly improbable
natural causes.[58]
Enlarge Image[59]
Kaita
draws a moral from this episode: ĹWe need always to keep in mind two separate
questions: how well does the evidence support design? And, Are we predisposed
to reject design apart from the evidence? The first is a scientific question.
The second is a philosophical question.ĺ[60] For those who wish to exclude the possibility of intelligent design a priori, Ĺthe motive is based not on ôjust the factsö but on philosophical prejudiceĺ.[61]
Such philosophical prejudice frequently results in question-begging and circular arguments. Philosopher of science Stephen C. Meyer calculates that: Ĺthe probability of constructing a rather short functional protein at random [is] so small as to be effectively zero. . .ĺ[62] If we ask Dawkins for the best explanation of the complex molecules of life he says: ĹNobody knows how it happened but, somehow, without violating the laws of physics and chemistry, a molecule arose that just happened to have the property of self-copying ľ a replicator.ĺ[63] If no one knows what happened, how does Dawkins know that what happened didnĺt Ĺviolate the laws of physics and chemistryĺ? He simply begs the question. Here, at least, is a frank admission of ignorance: ĹI would have to be more of a chemist that I am to know how likely it is that you are going to get such molecules,ĺ says Dawkins, ĹI donĺt know how difficult it would be to achieve that chemically.ĺ[64] Consider how significant it is to find Dawkins, who accuses theists of blind faith and who exhorts people to always ask, ĹWhat kind of evidence is there for that?ĺ,[65] believing in what he calls the Ĺsine qua nonĺ[66] of evolution without empirical evidence! Benjamin Wiker comments that Dawkinsĺ: Ĺlapse into an irrational faith in the powers of chance to avoid [intelligent design]. . . is not evidence itself but a telling lapse into a materialist credo quia absurdum estĺ.[67] Dawkins is not alone in making this sort of leap of faith. Harvard biologist George Wald dismissed a design explanation for life but candidly admitted:
I will not believe that
philosophically because I do not want to believe in God. Therefore, I choose to
believe in that which I know is scientifically impossible: spontaneous
generation [that life arose from non-living matter] arising to evolution.[68]
Phillip
E. Johnson complains: ĹThe naturalistic evolution of life from prebiotic
chemicals and its subsequent naturalistic evolution into complexity. . . is
assumed as a matter of first principle. . .ĺ[69]
As G. A. Kerkut of the Department of Physiology and Biochemistry at the
University of Southampton writes, it is: Ĺa matter of faith on the part of the
biologist that biogenesis did occur. . .ĺ[70]
For scientists like Dawkins and Wald, who reject an Ĺopenĺ philosophy of
science, the hypothesis that life Ĺjust happenedĺ is a philosophical
deduction entailed by the assumption of naturalism. To approach biology
without begging-the-question like this does not mean excluding consideration of
the theory of abiogenesis, or evolution (as Darwinians exclude consideration of
intelligent design). Rather, it means letting the evidence speak for itself and
following the evidence wherever it leads.[71]
Johnson observes that: ĹDarwinism is the answer to a specific question
that grows out of philosophical naturalism. . . The questions is: How must
creation have occurred if we assume that God [or other intelligent designer/s]
had nothing to do with it?ĺ[72]
Design theorists are more than happy to confirm that neo-Darwinism is the best
available answer to that question; but answering that question is not at
all the same thing as answering this far more interesting, unbiased and
important question: ĹHow did creation actually occur?ĺ
Scrabbling for the Right Explanation:
Understanding CSI
To understand what design theorists mean by CSI,
imagine a game of scrabble. A long string of random letters drawn from a
scrabble bag is complex without being specified - that is, without conforming
to a non ad hoc pattern that we havenĺt simply read off the object or
event in question. A short sequence of letters like Ĺthisĺ or Ĺthatĺ is
specified (it conforms to a non ad hoc, independent pattern) without
being sufficiently complex to outstrip the capacity of chance to explain this
conformity (letters drawn at random from the scrabble bag will occasionally
form a short word.) However, this article is both specified (conforming to the
independent functional requirements of grammatical English language use) and
sufficiently complex (doing so at a level of complexity that makes it
unreasonable to attribute this match to dumb luck) to trigger a design
inference. It would be unreasonable to suggest that I produced this article by
randomly drawing tiles from a scrabble bag!
Philosopher William Lane Craig explains the logic the
design inference with a simple illustration:
Bob is
given a new car for his birthday. There are millions of license plate numbers,
and it is therefore highly unlikely that Bob would get, say, CHT 4271. Yet that
plate on his birthday car would occasion no special interest. But suppose Bob,
who was born on 8 August 1949 finds BOB 8849 on the license plate of his
birthday car. He would be obtuse if he shrugged this off with the comment,
ĹWell, it had to have some license plate, and any number is equally
improbable. . .ĺ[73]
Bobĺs car having the first license plate was complex (unlikely) but
it wasnĺt noteworthy because it wasnĺt specified. Bobĺs car having the second
license plate was noteworthy because in addition to being complex it was also
specified (it matched the independently given pattern of Bobĺs name and
birth-date). Hence, according to William A. Dembski: Ĺgiven an event,
object, or structure, to convince ourselves that it is designed we need to show
that it is improbably (i.e. complex) and suitably patterned (i.e. specified).ĺ[74]
There
Just Arenĺt Enough Monkeys
áIt is
popularly thought that Ĺif you have enough monkeys banging randomly on
typewriters they will eventually type the works of William Shakespeareĺ, but
there is a real world limit to the number of Ĺmonkeysĺ available and the time
in which they can type. Indeed, I heard about a zoo that put a typewriter in
their monkey cage: the monkeys defecated in the typewriter and hit the same key
over and over again!
(Monkey picture from ĹMonkey Shakespeare Simulatorĺ[75])
 To take the point less literally, the online ĹMonkey Shakespeare
Simulatorĺ uses logged-on computers to simulate an ever-increasing population
of Ĺmonkeysĺ randomly typing (on simplified keyboards, at a rate of one dayĺs
typing per second), and compares the results to Shakespeareĺs works.[76]
As of October 2004, the best result was twenty Ĺlettersĺ (if one includes
spaces and full stops) from Coriolanus, after 462,060,000,000 billion
billion Ĺmonkey-yearsĺ![77] Getting better results is clearly
going to take a lot more monkey-years; but as Dembski would point out,
we canĺt simply keep giving ourselves free Ĺmonkey-yearsĺ. ĹJust where the
probabilities cutoff is can be debated,ĺ says Dembski, Ĺbut that there is a
probabilistic cutoff beyond which chance becomes an unacceptable explanation is
clear.ĺ[78]
As Richard Dawkins affirms: ĹWe can accept a certain amount of luck in our
explanations, but not too much.ĺ[79]
In fact, limiting the explanatory capacity of Ĺchanceĺ is crucial to the
integrity of science: ĹIf we allow ourselves too many ôwildcardö bits of
information. . . we can explain anything be reference to chance.ĺ[80] Allowing ourselves too
many Ĺwildcard bits of informationĺ commits the inflationary fallacy: Ĺthe
problem inherent in the inflationary fallacy is always that it multiplies
probabilistic resources in the absence of independent evidence that such
resources exist.ĺ[81]
To take the point less literally, the online ĹMonkey Shakespeare
Simulatorĺ uses logged-on computers to simulate an ever-increasing population
of Ĺmonkeysĺ randomly typing (on simplified keyboards, at a rate of one dayĺs
typing per second), and compares the results to Shakespeareĺs works.[76]
As of October 2004, the best result was twenty Ĺlettersĺ (if one includes
spaces and full stops) from Coriolanus, after 462,060,000,000 billion
billion Ĺmonkey-yearsĺ![77] Getting better results is clearly
going to take a lot more monkey-years; but as Dembski would point out,
we canĺt simply keep giving ourselves free Ĺmonkey-yearsĺ. ĹJust where the
probabilities cutoff is can be debated,ĺ says Dembski, Ĺbut that there is a
probabilistic cutoff beyond which chance becomes an unacceptable explanation is
clear.ĺ[78]
As Richard Dawkins affirms: ĹWe can accept a certain amount of luck in our
explanations, but not too much.ĺ[79]
In fact, limiting the explanatory capacity of Ĺchanceĺ is crucial to the
integrity of science: ĹIf we allow ourselves too many ôwildcardö bits of
information. . . we can explain anything be reference to chance.ĺ[80] Allowing ourselves too
many Ĺwildcard bits of informationĺ commits the inflationary fallacy: Ĺthe
problem inherent in the inflationary fallacy is always that it multiplies
probabilistic resources in the absence of independent evidence that such
resources exist.ĺ[81]
Postulating unlimited probabilistic resources makes it impossible to
warrant attributing anything to design.á Is Dawkins a good writer or does he
simply move his hands over his computer keyboard in the right way by luck?
Dawkinsĺ books could happen by chance, if we assume the existence of
sufficiently large probabilistic resources: ĹUnlimited probabilistic resources
make bizarre possibilities unavoidable on a grand scale.ĺ[82] Dembski calculates a
stringent probability bound of 10-150 based on the number of
elementary particles in the universe, the duration of the observable universe
and the Plank time.
Designed
or Designoid?
In Climbing
Mount Improbable Richard Dawkins draws a distinction between objects that
exhibit evidence of being designed and objects that give the superficial
impression that they exhibit evidence of being designed but on closer
inspection do not, which he calls Ĺdesignoidĺ.[83] Dawkins illustrates the
concept of being designoid with a hillside that suggests a human profile: ĹOnce
you have been told, you can just see a slight resemblance to either John or
Robert Kennedy. But some donĺt see it and it is certainly easy to believe that
the resemblance is accidental.ĺ[84]
Dawkins contrasts this Kennedy-esque hillside with the four presidentĺs heads
carved into Mt. Rushmore in America, which Ĺare obviously not accidental: they
have design written all over them.ĺ[85]
Although Dawkins defines biology as Ĺthe study of complicated things
that give the appearance of having been designed for a purposeĺ,[86] he believes that
appearances are deceiving. All biological things that appear to be designed are
designoid: ĹDesignoid objects look designed, so much so that some people
ľ probably, alas, most people ľ think that they are designed. These
people are wrong. . .á the true explanation ľ Darwinian natural selection ľ is
very different.ĺ[87]
Teasing apart Dawkinsĺ rhetoric is a rewarding hobby. His original
illustration of a designoid is of something that gives the superficial
impression that it exhibits evidence of being designed. People have to have the
resemblance between the hillside and Kennedy pointed out to them, some people
Ĺdonĺt see itĺ[88],
and Ĺit is certainly easy to believe that the resemblance is accidental.ĺ[89] However, Dawkins wants to
convince us that although some biological objects give such a strong appearance
of design that Ĺmost peopleĺ[90]
intuitively think that they are designed, they are merely designoid. On the
face of things, Dawkinsĺ design/designoid distinction actually supports
the majority opinion that life is the product of design. Some natural objects
are surely more analogous to Mt. Rushmore than they are to the Kenney-esque
hillside. In which case, how does Dawkins justify his confident, universal
negative claim (the hardest sort of claim to prove) that absolutely no
biological objects are designed, and that any biological object giving the
appearance, however strong, of design, is merely Ĺdesignoidĺ? Indeed, if
the paradigm Ĺdesignoidĺ object is the Kennedy-esque hillside, then such a
broad application of the concept is surely inappropriate. The meaning of
Dawkinsĺ flagship term shifts from Ĺthings that give the superficial impression
that they exhibit clear evidence of being designed, but on closer inspection do
notĺ, to Ĺthings that give every appearance of being designed, but are notĺ,
thereby exhibiting the logical fallacy of Ĺequivocationĺ.[91]
If Dawkinsĺ hypothesis is simply, as Alan Keith suggests, that Ĺsome
things that appear to be designed are not in fact designedĺ[92], what justification does
he give for thinking that life only appears to be designed?á Itĺs easy
enough to tell design and designoid apart as Dawkinsĺ introduces the
distinction; but how are we to tell design from designoid given his implicit
re-definition of these terms? We cannot! Dawkins applies the first distinction
on the basis of observational features of the objects in question, but he
applies the second distinction on the basis of his commitment to
metaphysical naturalism. A consistent, non-question-begging understanding
and application of Dawkinsĺ design/designoid distinction seems to support the
conclusion that life is the product of design, and hence of purpose.
Pigliucci
et al to the Rescue?
Reviewing this
criticism of the design/designoid argument[93],
Massimo Pigliucci[94]
and the members of his graduate class on evolutionary thinking at the
University of Tennessee, Knoxville respond that this objection to Dawkins Ĺis
partly rightĺ.[95]
According to Professor Pigliucci et al: ĹDawkins did choose a bad
example, and for fundamentally wrong reasonsĺ[96]
when he illustrated the concept of a Ĺdesignoidĺ with a hillside that looks a
bit like a Kennedy from the right angle. The problem with Dawkinsĺ analogy,
according to Pigliucci et al, is that: ĹThe resemblance of the cliff
outcropping to a human face is the result of entirely random causes. . . while
biological organisms are the outcome of two processes: mutation (which
is indeed random) and natural selection (which is anything but random).ĺ[97] Of course, whether or not
everything about all biological organisms is the outcome of nothing
but these natural forces, one random (mutation) and one non-random (natural
selection), is the very point at issue. Pigliucci et al agree with me
that ĹDawkinsĺ designoids donĺt cut itĺ, but argue that this is because
his original illustration provides a poor analogy for the organism producing
forces at work in evolution:
Dawkinsĺ fundamental
point can be rescued by simply using a better analogy.á There are natural,
non-biological, processes that convey the impression of intelligent design and
provide us with a more closer parallel to evolution. For example, on many rocky
beaches, pebbles are sorted by size going from the waterline towards the
interior, in a distinctly non-random pattern.á This is not because somebody got
all the pebbles out of the ocean, carefully weighed them, and then constructed
the beach.á Rather, the pattern was created by the joint action of two
processes: the (random) action of waves and the (non-random) effects of
gravity.[98]
Is this new
analogy for Ĺdesignoidĺ any better than Dawkinsĺ? No. If anything, it is a
worse analogy, because people donĺt intuit the presence of design when
observing beaches! The sorting of pebbles on a beach simply does not Ĺconvey
the impression of intelligent designĺ, whereas Dawkinsĺ Kennedy-esque hillside
does at least convey the initial impression of design. Hence Dawkinsĺ
hillside analogy, while it may be a worse analogy for the process of
evolution, is the better illustration of the concept of a Ĺdesignoidĺ, and the
pebble-sorting of Pigliucci et al, while it may be a better
analogy for evolution, is a worse illustration of the concept of a Ĺdesignoidĺ!
Like Dawkinsĺ, Pigliucci et al provide an illustration that tends to
reinforce the intuitive conclusion that the appearance of design in nature is not
deceptive.[99]
Philosopher Robert C. Koons observes
that: ĹThe Western philosophical tradition has bequeathed us two competing
metaphysical models: one in which everything is to be explained ultimately in
terms of blind and purposeless forces (the materialistic model); and one in
which purposefulness is a fundamental and irreducible reality (the teleological
model).ĺ[100] The most important question Ĺfrom an epistemological point of viewĺ[101], writes Koons, is this: Ĺwhere should
we locate the presumption of truth, and where the burden of proof?ĺ[102] Koons argues that:
There are compelling grounds for placing the burden of proof on the
materialistic model. Even stalwart Darwinists like Richard Dawkins admit that
the defining task of biology is to explain the existence of things that appear
to be designed. Cicero, in On the Nature of the Gods, Book II, reports
Aristotleĺs cave analogy: if a group of people had spent all of their lives
underground and then emerged on the surface, they would be bound to think of
the biologically rich world they discovered there to be the product of
intelligence. Only familiarity dulls our sense of wonder at the craftsmanship
of nature.
In his essay on the Intellectual
Powers of Man, eighteenth-century Scottish philosopher Thomas Reid cites
the capacity to recognize the signs of intelligent agency as part of the basic
equipment of the human mind. . . When this basic faculty of
intelligence-recognition considers the machinery of living things, the clear
answer it delivers is yes, there is intelligence and purposefulness displayed
in such machinery.[103]
While
Ĺthe natural deliverances of our sense of intelligence can be defeatedĺ[104], says Koons, Ĺthere is an
undeniable burden of proof that must first be assumed.ĺ[105] Question-begging
assertions like those made by Dawkins, Lewontin and Wald fail to defeat the
presumption of truth that should be accorded to belief in design.
Dawkinsĺ
Design Detection Criteria
Ironically
enough, Dawkinsĺ discussion of the design/designoid distinction actually
illustrates intelligent design methodology. Dawkins argues that while Ĺa rock
can weather into the shape of a nose seen from a certain vantage pointĺ,[106] [cf. figure 2] such a
rock is designoid (i.e. it is not clearly the produce of intelligent
design, because it is easy to believe that Ĺthe resemblance is accidental.ĺ[107]) Mt. Rushmore [cf. figure 1], on the other hand, is obviously not designoid: ĹIts four heads
are clearly designed,ĺ[108]
argues Dawkins, because: ĹThe sheer number of details [i.e. the amount of complexity]
in which the Mount Rushmore faces resemble the real things [i.e. the complexity
fits four independent specifications] is too great to have come about by
chance.ĺ[109]
In terms of mere possibility, says Dawkins: ĹThe weather could have done
the same job. . . But of all the possible ways of weathering a mountain,
only a tiny minority would be speaking likenesses of four particular human
beings.ĺ[110]
Hence: ĹEven if we didnĺt know the history of Mount Rushmore, weĺd estimate the
odds against its four heads being carved by accidental weathering as astronomically
high. . .ĺ[111]
 Figure 1. An intelligent design: Mount Rushmore with presidents Washington, Jefferson, Roosevelt, and Lincoln.
Figure 1. An intelligent design: Mount Rushmore with presidents Washington, Jefferson, Roosevelt, and Lincoln.
Figure 2. An accident of nature: President John F. Kennedyĺs profile formed by shadow cast by a large rock in Hawaii.[112]
In other
words, Mt. Rushmore is obviously the product of intelligent design because it
exhibits what information theorists call specified complexity, whereas
the hill-side is not obviously the product of intelligent design because it
does not exhibit specified complexity: Ĺit is certainly easy to believe
that the resemblance [i.e. the hillĺs conformation to a specification] is
accidental [i.e. is not complex].ĺ[113]
The significant point here is that Dawkinsĺ own design detection
criteria is obviously a pre-theoretic version of the design detection criteria
elaborated by William A. Dembski in such books as The Design Inference
(Cambridge, 1999) and No Free Lunch, (Rowman & Littlefield, 2001).
Mathematician & Philosopher Professor William A. Dembski has broken new ground in information theory with his work on the design inference from Ĺspecified complexityĺ. Among the most scholarly presentations of this work are: The Design Inference (Cambridge University Press, 1999), where Dembski explicates his Ĺexplanatory filterĺ & No Free Lunch (Rowman & Littlefield, 2002) which applies the insights of The Design Inference within biology.


From Pebbles on the Beach to Intelligent Design
Returning to the
beach example suggested by Pigliucci et al, let us ask the question:
ĹHow could pebbles on a beach convey clear evidence of intelligent
designĺ? A random spread of pebbles would not do the trick. Such a distribution
of pebbles may be complex, but it lacks specificity. Nor would having the
pebbles Ĺsorted by size going from the waterline towards the interior, in a
distinctly non-random patternĺ do the trick. Such a distribution of pebbles is
specific (Ĺdistinctly non-randomĺ) but not particularly complex. What
definitely would tip us off to the presence of design is if the pebbles
were arranged to spell out a message such as: ĹHello and welcome to the
seaside. We hope that you will enjoy this beach.ĺ Such an arrangement, which
exhibits an independent specification at very low probability, would obviously
imply intelligent design. Such an arrangement of matter not only Ĺlooks a bit
like it might be designed at first glanceĺ, like Dawkinsĺ hillside, but gives every
appearance of being designed. If one cannot infer design from such an
arrangement of matter, then matter simply cannot be arranged in a way that
clearly signals design! Only a dogmatic commitment to design-free explanations
would prevent one from saying that such a pattern was the result of design.
Design Without Equivocation
Itĺs all very
well to observe that some things that donĺt require explanation in terms of
design can give the superficial impression that they do require such an explanation
(e.g. Dawkinsĺ Kennedy-esque hillside), to call such things Ĺdesignoidĺ, and to
draw the moral that we should take care about saying that something requires
explanation in terms of design. It is quite another thing to observe Mt. Rushmore, or pebbles on a beach that spell ĹWe hope you enjoy this beachĺ, and to
assert that this pattern is Ĺdesignoidĺ! And yet, in effect, that is what
Dawkins does with the evidence for design.
However it is illustrated, the meaning of the key term in Dawkinsĺ
argument surreptitiously evolves under our noses. It begins life meaning
Ĺsomething that gives a superficial impression of designĺ and ends up meaning
Ĺa thing that gives every indication of being the product of design, but
isnĺt.ĺá Dawkins needs Ĺdesignoidĺ to carry this latter meaning because he
accepts that: ĹWhen a biologist looks at particular organs or organisms. . .
what he sees is a machine, which has every indication of being designed
for a purpose.ĺ[114]
However, such an equivocation turns a useful distinction into an empirically
vacuous concept that begs the question by smuggling in the metaphysical
assumption that intelligence is incapable of outperforming the design-producing
resources of nature in such a way as to leave reliable, empirically detectable indicators
of its activity. Yet when Dawkins is not concerned with design-proofing
biology, he holds that intelligence is capable of outperforming the
design-producing resources of nature in such a way as to leave reliable,
empirically detectable indicators of its activity. Indeed, he employs a version
of Dembskiĺs ĹDesign Filterĺ, motivating the inference to design in the
presence of specified complexity.
Only Positive Results
The ĹDesign
Filterĺ elaborated by Dawkins and Dembski is only a positive test for
design. Suppose an artist carefully sculpts a hillside by cleverly mimicking
natural weathering so as to produce a profile vaguely like that of a
man. The design filter would not detect the activity of intelligence in that
hillside. As far as the filter is concerned, the hillside is, at most, designoid.
It might be the product of design, it might superficially look
like objects that the filter would attribute to design, but the filter gives us
no reason to think that it is the product of design because the
hillside, while complex, is not specified. On the other hand, if the hillside
in question were carved into a detailed (i.e. specific) picture of four
American presidents, then the filter would detect design, because such a
pattern is both complex and specified. The filter canĺt rule design
out, but it can rule it in.
Part II: ID Theory Applied to Nature
Ĺintelligent design is an empirical hypothesis that
follows easily from the dataĺ ľ Michael J. Behe[115]
ID & Cosmology ľ Cracking Fine Tuning
The
basic physical laws of nature are Ĺfinely tunedĺ, or specified, in the sense
that if they were only a little different that they in fact are, then the
existence of intelligent life would be impossible. This is a fact that many
scientists and philosophers take to indicate design - even if they feel
uncomfortable with such an implication. As Stephen Hawking once admitted: ĹThe
odds against a universe like ours emerging out of something like the big bang
are enormous. . . I think clearly there are religious implications. . . But I
think most scientists prefer to shy away from the religious side of it.ĺ[116] Shying away from a
conclusion supported by evidence is hardly a scientific attitude to adopt.
According to Massimo Pigliucci: ĹShould we conclusively determine that the
probability of existence of our universe is infinitesimally small, and should
we fail to explain why physical constants have assumed the quantities that we
observe, the possibility of a designed universe would have to be considered
seriously.ĺ[117]
Hence, in discussing the fine-tuning of the cosmos, Pigliucci lays down what
amounts to a pre-theoretic version of Dembskiĺs explanatory filter, which
infers design when an independent specification (e.g. the set of physical laws
required by a life sustaining universe) is exhibited at sufficiently low
probability. Pigliucci and design theorists differ on whether we can infer that
our universe is the product of design, but at least there would appear to be
agreement on the criteria for making such a judgement.
Settling the question one-way or the other is a matter of evidence.
Richard Dawkins fails to make the connection between specified
complexity as a hallmark of intelligent design and the issue of cosmic
fine-tuning, but as has already been observed, he clearly accepts the same
design-detection criteria as Pigliucci and Dembski. Moreover, in establishing
CSI as a hallmark of intelligent design, Dawkins provides an excellent analogy
for understanding cosmic fine-tuning:á ĹOf all the unique and, with hindsight
equally improbable, positions of the combination lock,ĺ observes Dawkins, Ĺonly
one opens the lock. . . The uniqueness of the arrangement. . . that opens the
safe, [has] nothing to do with hindsight. It is specified in advance.ĺ[118] According to Dawkins, the
best explanation of an open safe is not that someone got lucky, but that
someone knew the specific and complex combination required to
open it. But isnĺt the Ĺfine-tuningĺ of the universe discovered by cosmologists
analogous to Dawkinsĺ example of a cracked combination lock?
Douglas Adams, author of The Hitchhikerĺs Guide to
the Galaxy, thought he had a knockdown response to the Ĺfine-tuningĺ
argument, likening it to a puddle of water arguing that, since the dip in the
ground it inhabited seemed to fit it so well, the dip must have been created
with its existence in mind: ĹThis is. . . an interesting hole I find myself in
ľ fits me rather neatly, doesnĺt it? In fact it fits me staggeringly well, must
have been made to have me in it!ĺ[119]
While this analogy makes for a nice piece of humour, as a rebuttal of the
fine-tuning argument it is deeply flawed. Water will fit any shape hole ľ the
fit between the hole and the water can be explained wholly by reference to the
nature of water. However, life will not fit just any old environment. The fit
between our cosmic environment and life cannot be explained wholly by reference
to the nature of life: ĹIn reviewing the physical laws and the numerical values
of fundamental constants, one encounters a remarkable precision in these values
such that only small changes in the fundamental constants. . . would yield a
universe without galaxies, starts, atoms or even nuclei, and consequently,
without the capacity for life.ĺ[120]
As Moreland and Craig note: ĹAn observer who has evolved within the universe
should regard it as highly probable that he will find the basic conditions of
the universe fine-tuned for his existence; but he should not infer that it is
therefore highly probable that such a fine-tuned universe exists at all.ĺ[121]
Moreland and Craig illustrate the sense in which the
fine-tuning of the universe is a highly improbable fact:
Take a
sheet of paper and place upon it a red dot. That dot represents our universe.
Now alter slightly one or more of the finely tuned constants and physical
quantities which have been the focus of our attention. As a result we have a
description of another universe, which we may represent by a new dot in the
proximity of the first. If that new set of constants and quantities describes a
life-permitting universe, make it a red dot; if it describes a universe that is
life prohibiting, make it a blue dot. Now repeat the procedure arbitrarily many
times until the sheet is filled with dots. One winds up with a sea of blue with
only a few pin-points of red.[122]
The hypothetical question of whether or not universes
are possible that have wholly different physical variables but are life
permitting is irrelevant to the fine-tuning argument. Imagine a fly resting on
a large, blank area on a wall:
A
single shot is fired, and the bullet strikes the fly. Now even if the rest of
the wall outside the blank area is covered with flies, such that a randomly
fired bullet would probably hit one, nevertheless it remains highly improbable
that a single, randomly fired bullet would strike the solitary fly within the
large, blank area. In the same way, we need only concern ourselves with the
universes represented on our sheet in order to determine the probability of the
existence of a life-permitting universe.[123]
In the case of Dawkinsĺ cracked combination lock what calls for a
teleological explanation is not merely that an event of small probability has
taken place (other sequences of number dials are equally improbable), but the
fact that this small probability event is specified (as the sequence
that opens the lock). Likewise, in the case of cosmic fine-tuning, what calls
out for explanation in terms of design is not merely that an event of small
probability has taken place (the existence of a particular set of physical
laws), but the fact that this event is specified (as the set necessary
for a life sustaining universe). Indeed, the fine-tuning of the universe is
like cracking a safe with multiple combination locks, one lock for each
cosmic parameter: The matter-anti-matter balance Ĺhad to be accurate to one
part in ten billion for the universe to arise.ĺ[124] The expansion rate of the
universe from the big bang had to be accurate to one part in 1060,
while the force of gravity itself required fine-tuning to one part in 1040.[125] If the strong nuclear
force were 2 percent weaker protons and neutrons wouldnĺt stick together. If it
were 0.3 percent stronger hydrogen (a crucial component of biological systems)
could not exist.[126]
Oxford physicist Roger Penrose calculated that the original phase-space
volume required such exact fine-tuning that the ĹCreatorĺs aim must have been
[precise] to an accuracy of one part in 1010(123).ĺ[127] Penrose was only speaking
poetically of the ĹCreatorĺs aimĺ, but this sort of data is in fact best
explained by the existence of a real Creator. Finely tuned improbabilities
compound one another until the overall improbability of cosmic fine-tuning
being a fluke becomes unimaginably high. Don N. Page[128] of the Institute for
Advanced Study in Princeton, N.J., calculates the odds against the formation of
our universe at one in 10,000,000,000124! As Fred Hoyle complained:
ĹA common sense interpretation of the facts suggests that a superintellect has
monkeyed with physics.ĺ[129]
Some cosmologists attempt to avoid this conclusion by positing the
existence of a large number of universes (perhaps an infinite number), each
with a different set of laws. However, what guarantees that
all these universes have different laws is a mystery. Stephen Clark
comments:
It is a
mark of desperation that some atheistical materialists have chosen to believe
in infinite arrays of universes. . . rather than believe instead that this
well-adapted world is founded on intelligence. . . explaining away this world
by saying that all worlds happen (which does not follow anyway merely
from there being, we fantasize, an infinite array of worlds) merely enlarges
the problem ľ and destroys the basis of all explanation (since we could not, on
those terms, be right to be surprised at anything, including Pratchettĺs Discworld).[130]
The Ĺmany worlds
hypothesisĺ commits the inflationary fallacy, of multiplying our
explanatory resources without an independent reason for doing so, on a grand
scale. The best explanation of cosmic fine-tuning is that an
intelligence took careful Ĺaimĺ with the purpose of producing a life-sustaining
universe. When we submit the scientific evidence of cosmic fine-tuning to the
design-detection criteria accepted by scientists like Dawkins and Pigliucci we
find that the evidence supports the hypothesis of intelligent design.
ID & Biology
Within biology,
Ĺintelligent design is a theory of biological origins and development.ĺ[131] Design theorists claim that
the current dominance of neo-Darwinian theory is a function of the pool of live
explanatory options being artificially restricted by an unjustified
methodological constraint, namely, the philosophical presupposition that
natural sciences should only explain things in terms of the inherent capacities
of nature (whether or not those capacities are themselves designed). Design
theorists argue for a more Ĺopen philosophy of scienceĺ, and insist that the
important question is not whether neo-Darwinism is the best explanation given
a ban on mentioning intelligent agency, but whether it is the best
explanation, full stop. Design theorists argue: Ĺthat once hypotheses
positing Intelligent Design are allowed into the pool of live options, then the
explanatory superiority of the neo-Darwinian theory is no longer apparent. On
the contrary, its deficiencies, particularly in the explanatory power of its
mechanisms of random mutation and natural selection, stand in stark relief.ĺ[132]
About Abiogenesis
When we examine the chemical basis of life we find highly complex
contingent arrangements of matter that are specified by their biological
functionality. According to Dawkins: ĹComplicated things have some
quality, specifiable in advance, that is highly unlikely to have been acquired
by random chance alone.á In the case of living things, the quality that is
specified in advance is. . . the ability to propagate genes in reproduction.ĺ[133] These
complex systems cannot be explained by Darwinĺs theory of evolution, for the
simple reason that they must exist before anything can evolve. As Dawkins says:
Ĺ[For evolution to occur] you need raw materials that can self-replicate. . .
The sine qua non [that without which]. . . is self-replication.ĺ[134] Pigliucci
et al correctly observe that Ĺwhat is needed for a naturalistic theory
of origins is that the first replicators were simple enough to originate
randomly.ĺ[135]
On the naturalistic hypothesis the first replicators have to be simple enough
to originate randomly because natural selection cannot explain the
origin of anything that is a precondition of natural selection ľ and
replicators are a pre-condition of natural selection.
In a telling letter to Philosophy
Now magazine (August/September 2004) Antony Flew discussed Ĺthe limits of
the negative theological implications of Darwinĺs Theory of Evolution by
Natural Selection.ĺ Quoting from Darwin, Flew notes that the theory of
evolution by natural selection does not account for the origin of life, and
observes that ĹProbably Darwin himself believed that life was miraculously
breathed into that primordial form of not always consistently reproducing life
by God. . .ĺ[136]á
Flew also notes that:
the evidential situation
of natural (as opposed to revealed) theology has been transformed in the more
than fifty years since Watson and Crick won the Nobel Prize for their discovery
of the double helix structure of DNA. It has become inordinately difficult even
to begin to think about constructing a naturalistic theory of the evolution of
that first reproducing organism.[137]
The idea that the pre-requisites of evolution could
simply Ĺariseĺ, perhaps from some Ĺwarm little pondĺ of chemicals as Darwin
hypothesized, is known as abiogenesis, from the Greek a
(without), bios (life) and ginomai (to form). The concept is
popularly known today under the rubric of the hypothetical Ĺprimal soupĺ, and
simply means the supposed naturalistic origin of life from non-life. For
example, Isaac Asimov confidently asserts: Ĺmolecules in the ocean grew
gradually more complicated until, eventually, some molecule was somehow formed
that could bring about the organization of simpler molecules into another
molecule just like itself. With that, life began. . . gradually evolving to the
present state of affairs . . .ĺ[138]
The way Asimov presents it, the naturalistic origin of life sounds like
established history. But as Flewĺs comments indicate, it isnĺt. A recent New
Scientist cover story on ĹThe 10 Biggest Mysteries of Lifeĺ (New
Scientist, 4th September 2004) included the question ĹHow did life begin?ĺ
as one of the Ĺbiggest unanswered questionsĺ in biology. As Alan Hayward says
of abiogenesis: ĹIt all sounds so simple, and so plausible ľ as long as you
know nothing about microbiology.ĺ[139]
Once upon a time in a warm little pond
. . .
The concept of abiogenesis was originally held by ancient Greek
thinkers such as Anaximander and Aristotle, and was revived in the
mid-twentieth century when Stanley Miller and Harold Urey recreated in the
laboratory what they believed to be an accurate representation of the early
earthĺs atmosphere, and managed (whilst mostly producing oils and tars) to
produce some amino acids by passing an electric spark through their mixture of
gases. If the extrapolation from this experiment to the viability of the
naturalistic origin of life from non-life were sound, one could still ask:
ĹWhat accounts for the existence of a ôprimal soupö with the correct Ĺrecipeĺ
for life?ĺá The answer, as Benjamin Wiker points out, would track back to the
finely tuned laws of nature, and hence track back to design: ĹSince biological
evolution depends on stellar evolution . . . the necessity of fine-tuning for
biological evolution has already been proven. Even now, Darwinism cannot claim
to be designer-free.ĺ[140]
A finely tuned Ĺprimal soupĺ suggests an intelligent ĹPrimal Cookĺ! However,
ĹThe ôprebiotic soup hypothesis,ö popularized by Millerĺs experiment, came
under withering criticism from chemists for ignoring the role of competing and
destructive cross-reactions. . . that would be expected in any hypothetical
ocean or pond.ĺ[141]
Moreover, ĹMiller and Ureyĺs experiment only works as long as oxygen is absent
and certain critical ratios of hydrogen and carbon dioxide are maintained.ĺ[142] As Dean
L. Overman explains: ĹThe presence of even a small amount of oxygen,
assiduously avoided in the laboratories of these experiments, would prevent the
formation of amino acids and nucleotides. . .ĺ[143] Of
course, if oxygen were not present, the molecules of life would have been
unprotected from deadly ultraviolet radiation: ĹWhat we have then is a sort of
ôCatch 22ö situation. If we have oxygen we have no organic compounds, but if we
donĺt have oxygen we have none either.ĺ[144] Scientists now think that oxygen was
present in the early earthĺs atmosphere, and that Ĺthe atmosphere of the early
earth was mostly made up of carbon dioxide and ammonia [meaning that the
Miller-Urey] experiment was not relevant to origin of life scenarios.ĺ[145] Hurbert
P. Yockey comments: ĹThe ôWarm little pondö scenario was invented ad hoc
as a materialistic reductionist explanation of the origin of life. It is
unsupported by any other evidence and it will remain ad hoc until such
evidence is found.ĺ[146]
These problems aside: ĹThe information filled
molecules of life are much more complex and structured than previously
thought.ĺ[147]
Denyse OĺLeary reports how: ĹDarwin thought that the world under the microscope
consisted of simple jellies and crystals that could easily form randomly. He
could not have been more wrong. It was only with the dawn of biochemistry in
the 1950ĺs ľ when scientists were able to look into cells deeply and in detail
ľ that they realized how wrong he was.ĺ[148] Nobel Prize-winning physiologist George
Beadle reports that DNA Ĺwas believed by many to be a rather monotonous polymer
built of four kinds of nucleotide units arranged in segments of four that were
repeated manifoldĺ.[149]
According to fellow Nobel winner Max Delbruck: Ĺit was believed that DNA was a stupid
substance.ĺ[150]
However, we now know that DNA is very far from being a monotonous polymer, but
is in fact an exceedingly complex and Ĺcleverĺ substance. The products of
tightly controlled and unrealistic laboratory experiments fall far short of the
complex protein molecules required for life: ĹMillerĺs optimism has now all but
evaporated, as experiments based on his model have failed to produce a number
of components essential to life.ĺ[151]
Varghese reports: ĹMiller acknowledges today that the real problem is making
polymers ľ like proteins ľ from simple chemicals like amino acids, something
that still eludes experimentalists.ĺ[152] Mark Shea remarks:
it has
always struck me as odd to point to the immense concentration of intellect,
will, technology and energy it has taken to do relatively small things in the
extremely specialized conditions of the lab (which nature allegedly did without
benefit of any of this) and argue that this product of white-hot focus of
ultra-controlling human intelligence is clear evidence that absolutely no
intelligence was involved in the production of the rest of the vastly more
complex life we see around us. Itĺs like taking years to build a tiny house of
cards and then using this feat to say, ĹThere! This accomplishment shows the
Capitol Dome was therefore obviously the product of a hurricane in a marble
quarry.ĺ[153]
As Chemist Jonathan Sarfati writes: Ĺthe very roots of the alleged
evolutionary tree are in very bad shapeĺ.[154]
 |
In A Case Against Accident and Self-Organization (Rowman & Littlefield, 2001) Dean L. Overman uses logical principles and mathematical calculations to answer the questions that have long perplexed biologists and astrophysicists: Is it mathematically possible that accidental processes caused the formation of the first form of living matter from non-living matter? Could accidental processes have caused the formation of a universe compatible with life? Are current self-organization scenarios for the formation of the first living matter plausible? |
Dawkinsĺ Credo Quia Absurdem Est
ĹToday it takes a great deal of faith to be an honest scientist who
is an atheist.ĺ - Walter L. Bradley[155]
What is the best explanation for the amazing molecules of life?
Dawkins offers the following: ĹNobody knows how it happened but, somehow,
without violating the laws of physics and chemistry, a molecule arose that just
happened to have the property of self-copying ľ a replicator.ĺ[156] Here,
at least, is a frank admission of ignorance: ĹI would have to be more of a
chemist that I am to know how likely it is that you are going to get such
molecules,ĺ says Dawkins, ĹI donĺt know how difficult it would be to achieve
that chemically.ĺ[157]
Can we consider for a moment how significant and amazing it is that Professor
Dawkins, who regularly accuses theists of blind faith and who exhorts people to
always ask, ĹWhat kind of evidence is there for that?ĺ,[158] should
believe in what he calls the sine qua non of evolution without a
shred of evidence?[159]
As Johnson complains: ĹThe naturalistic evolution of life from prebiotic
chemicals and its subsequent naturalistic evolution into complexity. . . is
assumed as a matter of first principle. . .ĺ[160] Benjamin Wiker comments that Dawkinsĺ
Ĺlapse into an irrational faith in the powers of chance to avoid [intelligent
design]. . . is not evidence itself but a telling lapse into a materialist
credo quia absurdum estĺ.[161]
In fact, there doesnĺt appear to be anything like sufficient evidence
for abiogenesis, for as Walter L. Bradley observes: Ĺthe origin of a
sophisticated system that is both rich in information and capable of
reproducing itself has absolutely stymied origin-of-life scientistsĺ.[162]
Not only does naturalistic science lack an explanation
of how life is supposed to have arrived on the cosmic scene, it actually
lacks any evidence that life Ĺjust happenedĺ. As G. A. Kerkut of the
Department of Physiology and Biochemistry at the University of Southampton
writes, it is Ĺa matter of faith on the part of the biologist that biogenesis
did occur. . .ĺ[163]
That abiogenesis Ĺjust happenedĺ, as Dawkinsĺ comments make clear, is a
philosophical deduction entailed by the assumption of naturalism. It is, as
chemist Robert Shapiro writes: Ĺmythology rather than scienceĺ.[164]
Manfred Schidlowsky argues that: Ĺthe very fact that
life sprang up on earth constitutes conclusive proof of a primary reducing
environment [i.e. one like the Miller-Urey experiment used] since the latter is
a necessary prerequisite for chemical evolution and spontaneous origin of
lifeĺ.[165]
But as Overman comments: ĹThis is a good example of. . . circular reasoning. .
. in which evidence is ignored in order to maintain a myth, and the conclusion
is set forth in the premise.ĺ[166]
While theists and agnostics have a healthy bias in favour of naturalistic
explanations when these are adequate, they cannot treat such a circular
deduction as the unquestionable and absolute certainty that it must be for
Dawkins. Rather, they will be open to following the evidence.
Stephen C. Meyer calculates that: Ĺthe probability of
constructing a rather short functional protein at random [is] so small as to be
effectively zero. . .ĺ[167]
In other words, not only does naturalistic science lack an explanation of how
the chemistry of life arose, or evidence to show that life Ĺjust
happenedĺ, it also flies in the face of evidence that life didnĺt Ĺjustĺ
happen! As Thaxton, Bradley and Olsen note: Ĺa slowly emerging line or boundary
has appeared which shows observationally the limits of what can be expected
from matter and energy left to themselvesĺ.[168]
Dawkins, who affirms that ĹWe can accept a
certain amount of luck in our explanations, but not too muchĺ[169], admits
that the chance origin of life theory Ĺmay seem like a big stroke of luckĺ;[170] but he
seeks to mitigate against this admission by saying: Ĺit had to happen only
once. . . it may have happened on only one planet out of a billion billion
planets in the universeĺ.[171]
But this is mere optimistic hand waving. The universe Ĺprobably contains no
more than one planet for every thousand starsĺ,[172] and Ĺit
is unlikely that there are many, if any, other earth-like planets in the
universeĺ[173]
able to sustain life.
Benjamin Wiker relates some of the finely tuned
conditions that permit life on earth:
Our sun
is not a typical star but is one of the 9 percent most massive stars in our
galaxy, and is also very stable. Further, the sun hits the Goldilocks mean for
life ľ neither too hot (like a blue or white star) nor too cold (like a red
star) ľ and its peak emission is right at the visible part of the
electromagnetic spectrum ľ the very, very thin band where not only vision is
possible but also photosynthesis. Earth just ôhappensö to have the right
combination of atmospheric gases to block out almost all the harmful radiation
on the electromagnetic spectrum but, strangely enough, opens like a window for
visible light. Jupiter is deftly placed and sized so that it not only helps to
balance the Earthĺs orbit but also acts as a kind of debris magnet keeping
Earth from being pummeled. Our moon is just the right
size and distance to stabilize earthĺs axial tilt so that we have seasonal
variations but not wildly swinging temperature changes.[174]
Astronomer Hugh Ross lists 200 parameters required for
a life-bearing planet. Comparing the chances of a planet falling within these
parameters by chance alone with our best estimate of the total number of
planets in the universe (1022) he estimates that there is Ĺless than
1 chance in 10215ĺ of even one habitable planet existing in the
universe Ĺwithout invoking divine miraclesĺ.[175] Elsewhere, Ross writes that: Ĺfewer than a
trillionth of a trillionth of a percent of all stars will have a planet capable
of sustaining advanced life. Considering that the observable universe contains
less than a trillion galaxies, each averaging a hundred billion stars, we can
see that not even one planet would be expected, by natural processes alone, to
possess the necessary conditions to sustain life.ĺ[176]
Astrobiologists Peter D. Ward and Donald Brownlee conclude that: ĹIf some
god-like being could be given the opportunity to plan a sequence of events with
the express goal of duplicating our ôGarden of Edenö, that power would face a formidable
task. With the best intentions, but limited by natural laws and materials,
it is unlikely that Earth could ever be truly replicated.ĺ[177]
 |
In Rare Earth: Why Complex Life is Uncommon in the Universe (Springer-Verlag, 2000), palaeontologist Peter Ward and astronomer Donald Brownlee argue that complex life is rare in the universe and make a convincing case against the principle of mediocrity (i.e. ĹEarth isnĺt that specialĺ) that has ruled astronomy since Copernicus. |
 |
In The Privileged Planet (Regnery Publishing, 2004), astronomer Guillermo Gonzalez (whose research on the ĹGalactic Habitable Zoneĺ captured the October 2001 cover story of Scientific American) and philosopher Jay Richards use similar ĹRare Earthĺ data to Ward & Brownlee to argue that our planet is exquisitely fit not only to support life, but also gives us the best view of the universe, as if Earth-and the universe itself-were designed both for life and for scientific discovery. |
For another thing, according to Meyer, to generate a single
functional protein of 150 amino acids exceeds: Ĺ1 chance in 10180,
well beyond the most conservative estimates for the small probability bound. .
. it is extremely unlikely that a random search through all the possible amino
acid sequences could generate even a single relatively short functional protein
in the time available since the beginning of the universe. . .ĺ[178] In The
Blind Watchmaker, Dawkins simply says: ĹGiven enough time, anything is
possible.ĺ[179]
But there simply isnĺt enough time available to sustain the plausibility
of abiogenesis. Professor of Mathematics at Cardiff University, Chandra
Wickramasinghe, concludes: ĹLiving systems could not have been generated by
random processes, within a finite time-scale, in a finite universe.ĺ[180]
Dawkinsĺ blithe, hand waving ignorance of the evidence against the
naturalistic origin of life from non-life surely shows, as Michael Behe
comments, Ĺthe need to treat Darwinian scenarios. . . with a hermeneutic of
suspicionĺ.[181]
Behe goes on to observe: ĹSome scientists believe so strongly in Darwinism that
their critical judgments are affected, and they will unconsciously overlook
pretty obvious problems with Darwinian scenarios, or confidently assert things
which are objectively untrue.ĺ[182]
The evidence seems to point in quite another direction. Keith Ward takes stock
of the implications of the improbabilities of life Ĺjust happeningĺ: ĹIt seems
hugely improbable that, in the primeval seas of the planet earth, amino acids
should meet and combine to form large molecular structures capable of
self-replication. . . The motive for positing some sort of intelligent design
is almost overwhelming.ĺ[183]
Likewise,
Hoyle and Wickramasinghe conclude:
the
enormous information content of even the simplest living systems. . . cannot in
our view be generated by what are often called Ĺnaturalĺ processes. . . There
is no way in which we can expect to avoid the need for information, no way in
which we can simply get by with a bigger and better organic soup, as we
ourselves hoped might be possible. . . The correct position we think is. . . an
intelligence, which designed the biochemicals and gave rise to the origin of
carbonaceous life. . . This is tantamount to arguing that carbonaceous life was
invented by noncarbonaceous intelligence. . .[184]
Hoyle and Wickramasinghe stop short of making the philosophical
identification of their Ĺnoncarbonaceous intelligenceĺ with God, but note:
It is
ironic that the scientific facts throw Darwin out, but leave William Paley, a
figure of fun to the scientific world for more than a century, still in the
tournament with a chance of being the ultimate winner. . . Indeed, such a
theory is so obvious that one wonders why it is not widely accepted as being
self-evident. The reasons are psychological rather than scientific.[185]
Dawkins,
Design & DNA
Dembskiĺs
explicit formulation of the criteria for design detection used by Dawkins
naturally leads us to agree with Dawkins about Mt. Rushmore being designed.
However, if we apply the very same design-detection criteria to DNA, for
example, we get the very same conclusion: Design. Yet philosopher Norman L.
Geisler follows the evidence while Dawkins will not:
suppose I come upon a
round stratisfied stone and were asked how it came to be such. I might
plausibly answer that it was once laid down by water in layers which later
solidified by chemical action. One day it broke from a larger section of rock
and was subsequently rounded by the natural erosional processes of tumbling in
water. Suppose then. . . I come upon Mount Rushmore. . . Even if I knew nothing
about the origin of the faces, would I not come immediately to believe it was
an intelligent production and not the result of natural processes of erosion?
Yet why should a natural cause serve for the stone but not for the faces? For
this reason, namely, that when we come to inspect the faces on the mountain we
perceive ľ what we could not discover in the stone ľ that. . . they convey
specifically complex information. . .[186]
Thus far
Dawkins agrees, and for the same reasons; but he begs-the-question in order to
avoid extending that agreement to Geislerĺs consistent conclusion:
Suppose also that in
studying the genetic structure of a living organism, we discover that its DNA
has a highly complicated and unique information code, distinguished by its
specified complexity. . . would we not conclude that it most probably took
intelligence to produce a living organism?[187]
To
substitute the terms in Dawkinsĺ own argument: undirected natural causes could
have done the same job. But of all the possible ways of arranging amino
acids, only a tiny minority would match the biological specification for
functionality. Hence, even without knowing the history of DNA, weĺd
estimate the odds against its occurrence by natural processes as astronomically
high. But if natural processes didnĺt do the job, what did? Philosopher of
science Stephen C. Meyer has the answer:
Our experience-based
knowledge of information-flow confirms that systems with large amounts of
specified complexity (especially codes and languages) invariably originate from
an intelligent source. . . As Quastler (1964) put it, the Ĺcreation of new
information is habitually associated with conscious activity.ĺ (p. 16).
Experience teaches this obvious truth.[188]
The evidence
argues for intelligent design, but Dawkins allows the philosophical dogma of
naturalism to trump the evidence and shoehorn it into the naturalistically
acceptable category of Ĺdesignoidĺ, even though doing so requires an ad hoc
redefinition of his own term that ignores his own design detection criteria.
Darwinĺs Core Argument
According to
Professor Richard Norman, Darwinĺs core argument goes like this: Variations
exist within existing populations of domesticated plants and animals. Human
breeders have intelligently selected which set of characteristics get to breed,
such that Ĺsmall variations can be accumulated over many generations to
produce, say, a new breed of sheep of a new variety of rose.ĺ[189] Darwin argued that an
analogous process of selection happens in nature: Ĺwhat he calls ôthe struggle
for existenceö functions as a mechanism of selection comparable to selection
and breeding by human beings.ĺ[190]
Darwinĺs analogy between intelligent and natural selection implies
that a non-intelligent process is analogous to a process requiring the
involvement of intelligent agents, that a process in which the offspring are
exposed to harsh environmental conditions is analogous to a process in which
offspring are protected from the environment, and that a process in which
offspring can interbreed in the wild is analogous to a process where the
breeding of offspring is carefully orchestrated. This strains credulity in
itself.
Since there is Ĺno clear divideĺ between varieties and species, the
gradual accumulation of modifications can presumably produce changes that are
not just new varieties, but new species, etc. Hence, Ĺcontinued over vast
periods of timeĺ, this process of natural selection Ĺis sufficient to account
for the gradual emergence of all the species of living things which have
existedĺ.[191]
As Carl Zimmer asserts: ĹIf you accept microevolution, you get macroevolution
for free.ĺ[192]
Assessing Darwinĺs Core Argument
To quote
Professor Normanĺs summary of Darwinĺs argument:
- ĹWe know that
artificial [intelligent] selection of domesticated plants and animals can
produce new varieties.ĺ
- ĹWe know that
an analogous process of natural selection takes place in nature to produce
new varieties better adapted to their environment.ĺ
- ĹThere is no
reason why the process of natural selection which produces new varieties
may not, over sufficiently long periods of time, also produce varieties so
different as to constitute new species.ĺ
- ĹThe mechanism
of natural selection can therefore [i.e. may] explain how new species have
come into existence with features adapted to their environment.ĺ [193]
The argument
appears to be logically valid. Premise one is above reproach; as is premise two
(in that we know changing environmental conditions can alter the proportional
representation of pre-existing variations within a population of organisms).
However, premise three (which really needs to go further than the level of species,
and I will take this extension as read in order to avoid attacking a straw-man)
is a very large and therefore inherently shaky extrapolation from premise two.
As mathematician David Berlinski writes:
The most ardent creationist now accept micro-evolution as genuinely
Darwinian events. They had better: such are the facts. But the grand
evolutionary progressions, such as the transformation of a fish into a man, are
examples of macro-evolution. They remain out of reach, accessible only
at the end of an inferential trail.[194]
The further one
extends an inferential trail, the less solid it becomes. Our knowledge of
Ĺmicro-evolutionaryĺ changes does little to support belief in full-blown
Ĺmacro-evolutionĺ (evolution beyond the level of species, and/or the evolution
of new organs and body-plans). For example, after a 1977 drought in the Galapagos islands, scientists found that the surviving finches had beaks that were 4%
longer and 6% deeper than the average pre-drought beak. Then, after a period of
very wet conditions, scientists found that the average beak size of the
surviving finches was about 1% narrower than before the drought! This is a good
example of Ĺmicro-evolutionaryĺ change: ĹThe change in average beak size is an
example of minor variations being selected from genetic information already
present in the gene pool.ĺ[195]á
(Note that the change was cyclical and not in a uniform direction.) To
extrapolate from such examples of minor variations within a pre-existing
gene pool to the belief that ĹThere is no reason why the process of natural
selection which produces new varieties may not, over sufficiently long periods
of time, also produce varieties so different as to constitute new species [and
more]ĺ, is rather like arguing that since an Olympic runner can cover a mile in
four minutes there is no reason that he may not cover sixteen miles in
sixty-four minutes!á Obviously not. There are limiting factors that mean this
extrapolation is unrealistic:
Think of an archer shooting an arrow. Letĺs say that the arrow
travels at about 150 MPH. So, in half an hour, it should be able to hit a
target 75 miles away, right? Obviously, that wonĺt happen. There are limiting
forces like friction and gravity that dramatically slow the speed of the arrow after
the first 50 yards or so.[196]
Simply assuming
that there are no limiting factors does not prove that there are none! There is
reason to believe that evolution might account for speciation (this
hypothesis is the smallest and hence most secure extrapolation from the
evidence of observed micro-evolution, and there is some circumstantial evidence
in its favour[197]).
However, evidence for more extensive macro-evolutionary change, especially
change involving the appearance of new organs and/or new body plans, is
lacking. And as David DeWolf, Stephen C. Meyer and Mark E. DeForrest observe:
Evidence from developmental biology suggests clear limits to the
amount of evolutionary change that organisms can undergo, [hence there is
evidence of limiting factors] casting doubt on the Darwinian theory of common
descent and suggesting a reason for morphological stasis in the fossil record.[198]
Skeletons in the Fossil Record
ĹAs an illustration of the fossil record, the Tree of Life is a dismal failure. But it
is a good representation of Darwinĺs theory.ĺ ľ Jonathan Wells[199]
The fossil
record does little to endorse Darwinian gradualism, as Robert C. Koons
observes:
the fossil record of the family tree of evolution
is so gappy that it consists of a great deal more gap than tree. This is
especially true where the record is most complete, as in the case of the
invertebrates. The missing links that have been found, like the Archaeopteryx
or Australopithecus, are better described as mosaics: re-combinations of
adaptations found in what are assumed to be related families. Given that the
forms of life found in the fossil record are more numerous and variegated than
those we find alive today, it is not at all surprising that we should find
fossil forms that are Ĺintermediateĺ in some vague sense between living forms.
What we donĺt find is the kind of continuous, seamless web of transformation of
adaptive structures that would be needed to confirm the truth of Darwinism.[200]
Robert F. Dehaan
and John L. Wiester report that: Ĺeach phylum is self-bounded.á Indeed, there
are no transitional forms between them, as predicted by Darwinian theory.ĺ[201] David B. Kitts writes that:
Despite the bright promise that paleontology provides a means of
seeing evolution, it has presented some nasty difficulties for evolutionists,
the most notorious of which is the presence of Ĺgapsĺ in the fossil record.
Evolution requires intermediate forms between species and paleontology does not
provide them.[202]
Kitts concludes:
ĹThe gaps must therefore be a contingent feature of the recordĺ[203], but this is to argue from
the assumption of macroevolution to an ad hoc explanation for the poor
fit between theory and evidence, and to accept the fact that the fossil
record fails to verify Darwinĺs theory. The fossil record has become
something for evolutionary theory to explain away rather than something that
provides positive support for macro-evolution.
Moreover, according to Luther Sunderland: ĹThe gaps between major
groups of organisms have been growing ever wider and more undeniable. They can
no longer be ignored or rationalized away with appeals to the imperfection of
the fossil record.ĺ[204]
With around 250 million catalogued fossils, representing some 250,000 species,
Ĺthe problem does certainly not appear to be an imperfect record. Many
scientists have conceded that the fossil record is sufficiently complete to
provide an accurate portrait of the geologic record.ĺ[205] For example, Niles Eldridge
writes: ĹThe record jumps, and all the evidence shows that the record is real:
the gaps we see reflect real events in lifeĺs history ľ not the artefact of a
poor fossil record.ĺ[206]
University of Chicago Professor of geology David Raup says:
we are now about 120 years after Darwin and the knowledge of the
fossil records has been greatly expanded. We now have a quarter of a million
fossil species but the situation hasnĺt changed much. The record of evolution
is still surprisingly jerky and, ironically, we have even fewer examples of
evolutionary transition than we had in Darwinĺs time.[207]
A study
published in the February 26th 1999 issue of Science
indicates that Ĺthe fossil record is virtually complete in what it has to
reveal.ĺ[208]
The study combined data analysis of hundreds of mammal fossils with a
mathematical model of evolutionary branching patterns in an attempt to
determine the completeness of the fossil record prior to 65 million years ago:
The researchers concluded that the fossil preservation rate is high
ľ high enough that the probability that modern mammals existed more than 65
million years ago without leaving fossils is just .2 percent (two tenths of one
percent). Study author Christine Janis, professor of ecology and evolutionary
biology at Brown University, proclaimed, ĹThe fossil record for that period is
good enough for us to say that those species would most likely have been
preserved if they had been there.ĺ[209]
Phillip E.
Johnson summarises the fossil situation:
the fossil evidence is very difficult to reconcile with the
Darwinist scenario. If all living species descended from common ancestors by an
accumulation of tiny steps, then there once must have existed a veritable
universe of transitional intermediate forms linking the vastly different
organisms of today, such as moths, trees, and humans, with their hypothetical
common ancestors. From Darwinĺs time to the present, palaeontologists have
hoped to find the ancestors and transitional intermediates and trace the course
of macroevolution. Despite claims of success in some areas, however, the
results have been on the whole disappointing. That the fossil record is in
important respects hostile to a Darwinist interpretation has long been known to
insiders as the Ĺtrade secret of palaeontology.ĺ The secret is now coming out
into the open.[210]
The fossil
record not only fails to support Darwinism, it actually contradicts Darwinism.
This particular Darwinian skeleton is out of the fossil cupboard! As Johnsonĺs
comments indicate, the evidence is not uniformly inimical to macro-evolution,
but even in those cases where evolutionists can claim to find evidence for
macro-evolution, things are not open-and-shut. Consider Niles Eldridgeĺs
comments, made in the context of the famous Ĺhorse sequenceĺ of fossils, where
it turns out that Ĺthe species that were supposed to align in an evolutionary
lineage actually persist unchanged and co-exist in the fossil recordĺ[211]:
There have been an awful lot of stories, some more imaginative than
others, about what the nature of that history [of life] really is. The most
famous example. . . is the exhibition on horse evolution prepared perhaps fifty
years ago. That has been presented as the literal truth in text-book after
text-book.á Now I think that this is lamentable, particularly when people who
propose those kinds of stories may themselves be aware of the speculative
nature of some of that stuff.[212]
Hence, while some fossil evidence might be made
to fit the assumption of macroevolution, when taken as a whole, the fossil
evidence can hardly be said to support turning that assumption into a
conclusion.
Steve M. Stanley, professor of paleobiology at John Hopkins
University, confirms that ĹThe known fossil record fails to document a single
example of phylitic evolution accomplishing a major morphological transition
and hence offers no evidence that the gradualistic model can be valid.ĺ[213] Gareth Nelson, a senior
zoologist at the American Museum of Natural History, likewise admits that
Ĺevidence, or proof, of origins. . . of all the major groups of life, of all
the minor groups of life, indeed of all the species ľ is weak or nonexistent
when measured on an absolute scale.ĺ[214]
Robert Carroll observes that as our knowledge of the fossil record has
increased over the past century is has only emphasised: Ĺhow wrong Darwin was in extrapolating the pattern of long-term evolution from that observed within
populations and species.ĺ[215]á
Indeed, Mark Hartwig points out that:
According to Darwinism, new phyla are produced by the gradual
divergence of species. As species split off from each other over time, they
eventually become so dissimilar as to constitute a whole new body plan.
Therefore, we should see new species slowly appearing over time, followed by
the much slower appearance of new phyla ľ what Harvard palaeontologist Stephen
Jay Gould called a Ĺcone of increasing diversity.ĺ Instead, the cone is upside
down.[216]
Not only is Darwinĺs extrapolation inherently risky given its small evidential base, the fossil record
contradicts it. Nowhere is this more apparent than in the ĹCambrian explosionĺ.
The Cambrian Explosion and Intelligent Design
ĹDarwinism
cannot explain the Cambrian explosion. . . we need a new theory.ĺ ľ Dr.
Jun-Yuan Chen, palaeontologist, Nanking Institute of Geology.[217]
The
contradiction between Darwinian theory and the fossil evidence is hardly
surprising when one considers the fact that: Ĺempirically derived estimates of
mutation rates in extant organisms suggest that the kind of large-scale
morphological changes that occurred in the Cambrian would have required far
more time than the duration of the explosion. . .ĺ[218] Francis J. Beckwith reports
how:
evolutionists admit that the record does not reveal gradual
development from simple to more complex species, as predicted by Darwin. Rather, in what is called the ĹCambrian explosionĺ the record reveals the sudden
appearance at differing times of information-rich organisms within a
hierarchical diversity of species with apparently no precursors. Their body
plans with their improbable arrangement of parts including the information
content of their DNA. . . exhibit the characteristics of specified complexity,
intelligent design. Hence, some design theorists employ the facts of the
Cambrian explosion in their arguments for ID. . .[219]
For example, in
a paper on ĹThe Origin of Biological Information and The Higher Taxonomic
Categoriesĺ, published in the Proceedings of the Biological Society of
Washington (volume 117, no. 2, pp. 213-239), a peer-reviewed biology
journal published at the National Museum of Natural History at the Smithsonian
Institution in Washington D.C.[220],
Dr. Stephen C. Meyer treats Ĺthe problem of the origination of the higher
taxonomic groups as a manifestation of a deeper problem, namely, the problem of
the origin of the information. . . necessary to generate morphological
novelty.ĺ Meyer observes that an animal Ĺformĺ represents: Ĺa highly specific
and constrained arrangement of material components (among a much larger set of
possible arrangements).ĺ This understanding of Ĺformĺ reveals a link with Ĺthe
notion of information in its most theoretically general sense. . . producing
organismal form by definition requires the generation of information.ĺ Applying
these insights to the Cambrian explosion, Meyer notes that:
The Cambrian explosion represents a remarkable jump in the specified
complexity or Ĺcomplex specified informationĺ (CSI) of the biological world
[because] Functionally more complex animals [as appeared in the Cambrian]
require more cell types to perform their more diverse functions. New cell types
require many new and specialized proteins. New proteins, in turn, require new
genetic information. Thus an increase in the number of cell types implies (at
minimum) a considerable increase in the amount of specified genetic
information.
Studies of
modern animals suggests the sponges which appeared in the late Precambrian
would have required just five cell types: Ĺwhereas the more complex animals
that appeared in the Cambrian (e.g. arthropods) would have required fifty or
more cell types.ĺ Meyer argues that:
whether one envisions the evolutionary process beginning with a
non-coding region of the genome or a pre-existing functional gene, the
functional specificity and complexity of proteins impose some very stringent
limitations on the efficacy of mutation and selection. In the first case,
function must arise first, before natural selection can act to favor a novel
variation. In the second case, function must be continuously maintained in
order to prevent deleterious (or lethal) consequences to the organisms and to
allow further evolution. Yet the complexity and functional specificity of
proteins implies that both these conditions will be extremely difficult to
meet. Therefore, the neo-Darwinian mechanism appears to be inadequate to
generate the new information present in the novel genes and proteins that arise
with the Cambrian animals.
Moreover:
Mutations in genes that are expressed late in the development of an
organism will not affect the body plan. . . Thus, events expressed early in the
development of organisms have the only realistic chance of producing large-scale
macroevolutionary change. . . Yet recent studies in developmental biology make
clear that mutations expressed early in development typically have deleterious
effects.
This is because:
Ĺprocesses of development are tightly integrated spatially and temporally such
that changes early in development will require a host of other coordinated
changes in separate but functionally interrelated developmental processes
downstream.ĺ J.F. McDonald has called this problem Ĺa great Darwinian paradoxĺ:[221]
McDonald notes that genes that are observed to vary within natural
populations do not lead to major adaptive changes, while genes that could cause
major changes ľ the very stuff of macroevolution ľ apparently do not vary. In
other words, mutations of a kind that macroevolution doesnĺt need (namely,
viable genetic mutations in DNA expressed late in development) do occur, but
those that it does need (namely, beneficial body plan mutations expressed early
in development) apparently donĺt occur. According to Darwin natural selection
cannot act until favourable variations arise in a population. Yet there is no
evidence from developmental genetics that the kind of variations required by
neo-Darwinism ľ namely, favourable body plan mutations ľ ever occur.
Given the
inadequacy of evolutionary explanations of the CSI increase represented in the
Cambrian explosion, and given our every-day experience of intelligent agents as
sufficient causal explanations for such increases in CSI, Meyer concludes that:
Ĺexperience-based analysis of the causal powers of various explanatory
hypotheses suggests purposive or intelligent design as a causally adequate ľ
and perhaps the most causally adequate ľ explanation for the origin of the
complex specified information required to build the Cambrian animals and the
novel forms the represent.ĺ
In their paper ĹThe Cambrian Explosion: Biologyĺs Big Bangĺ[222], Meyer, Ross, Nelson and
Chien observe that:
When we compare the pattern of fossilization in the actual fossil
record to the expected pattern given the neo-Darwinian mechanism, we encounter
significant dissonance. Neither the pace nor the mode of evolutionary change
match neo-Darwinian expectations. Indeed, the neo-Darwinism mechanism cannot
explain the geologically sudden origin or the major body plans to which the
term Ĺthe Cambrian explosionĺ principally refers. Further, the absence of
plausible transitional organisms, the pattern of disparity preceding diversity,
and the pattern of phyla first appearance all run counter to the neo-Darwinian
expectations. Although. . . the newer punctuationist model of evolutionary
change appears more consonant with some aspects of the Cambrian/Precambrian
fossil record, it too, fails to account for the extreme absence of transitional
intermediaries, the top-down pattern of diversity, and the pattern of phylum
first appearance. . .[223]
They note how
ĹThese problems underscore a more significant theoretical difficulty for
evolutionary theory generally, namely, the insufficiency of attempts to
extrapolate microevolutionary mechanisms to explain macroevolutionary
developmentĺ[224],
and go on to argue that:
we see in the fossil record several distinctive features or
hallmarks of designed systems, including: (1) a quantum or discontinuous
increase in specified complexity or information; (2) a top-down pattern of
innovation in which large-scale morphological disparity arises before
small-scale diversity; (3) the persistence of structural (or Ĺmorphologicalĺ)
disparities between separate organized systems; and (4) the discrete or
simultaneous emergence of functionally integrated parts within novel
organizational body plans. . .á in other words, intelligent design constitutes
the best, most causally adequate, explanation of the specific features of the
Cambrian explosion. . .[225]
The Extrapolation that Wonĺt Fly
ĹI have no
theological problem with random mutation as the vehicle of evolution; it still
leaves open the need for a source of nature and its laws. Similarly I have no
problem with the existence of unicorns. I just find both beliefs to be equally
fanciful.ĺ ľ Roy Abraham Varghese[226]
Phillip E.
Johnson argues that none of the examples of microevolution: Ĺprovides any
persuasive reason for believing that natural selection can produce new species,
new organs, or other major changes. . .ĺ[227]
Darwinĺs bold extrapolation from observed micro-evolutionary change to the
hypothesis that macroevolution accounts for all organic variation simply
assumed that there are no limiting factors analogous to the limiting
factors that came into play in the story about the Olympic runner or the story
about the archer; hence Professor Norman is building a house on sand when he
asserts that: Ĺthe Darwinian explanation shows how . . . these mechanisms can
account for the emergence of species [and more]ĺ.[228] By way of contrast, biologist
Keith Thompson of Oxford University says that: Ĺno one has satisfactorily
demonstrated a mechanism at the population genetic level by which
innumerable very small. . . changes could accumulate rapidly to produce large
changes: a process for the origin of the magnificently improbable from the
ineffably trivialĺ.[229]
At best, the Darwinian explanation suggests how the observed
mechanism of natural selection might account for the emergence of
species (and more) if there are no limiting factors that invalidate what
is at any rate a risky extrapolation from the available data. As David L. Hull
of the University of Wisconsin observes: ĹThe leading philosophers contemporary
with Darwin, John Herchet, William Whewell, and John Stewart Mill, were equally
adamant in their conviction that the Origin of Species was just one mass
of conjecture.ĺ[230]
They were right. As Nancy Pearcey and Charles Colson comment: ĹIt was a bold
speculation, but no one should be misled into thinking it was more than that. .
.á It is a conjecture, an extrapolation going far beyond any observed facts.ĺ[231]
Darwinĺs Ĺriskyĺ hypothesis (to borrow a term from philosopher of
science Karl Popper) was a good scientific hypothesis that was well worth
checking out, but 150 years of checking have indicated that Darwin was at best
only partly right: ĹYes, small-scale evolution is a factĺ, writes molecular
biophysicist Cornelius G. Hunter, Ĺbut there is no reason to think it is
unbounded. In fact, all our data suggests that small-scale evolution cannot
produce the sort of large-scale change Darwinism requires.ĺ[232] On the basis of the fossil
record, Dehaan and Wiester conclude: ĹThe Darwinian mechanism of selection and
variation does provide a plausible explanation for minor variations among
species, such as the variations of shapes in finchesĺ beaks. But this mechanism
plays no discernable part in the formation of major innovationsĺ[233] As Roger Lewin stated in his
summary of the Chicago ĹMacroevolutionĺ conference in 1980:
The central question of the Chicago conference was whether the
mechanisms underlying microevolution can be extrapolated to explain the
phenomena of macroevolution. At risk of doing violence to the position of some
people at this meeting, the answer can be given as a clear, No.[234]
Today, Cornelius
G. Hunter reports that:
if one looks in the research journals, one finds that evolutionists
are unsure whether small-scale evolution could possibly account for the needed
large-scale change. . . From genetics to palaeontology and other disciplines,
the message is that evolutionĺs necessary large-scale change does not appear to
be a simple case of small-scale change extrapolated over time.[235]
Hence Jonathan
Wellĺs argues that:
common ancestry is certainly true at the species level, but is it
true at higher levels? It becomes an increasingly uncertain inference the
higher we go in the taxonomic hierarchy. When you get to the level of phyla,
the major animal groups, itĺs a very, very shaky hypothesis. In fact, I would
say itĺs disconfirmed. The evidence just doesnĺt support it.[236]
The extrapolation from observed microevolution to an explanatorily
sufficient macroevolution (based on the assumption that there are no
limiting factors) is contradicted by our knowledge of mutation rates, developmental
biology, and the fossil record. Macro-evolution just doesnĺt seemed to have
happened. As Johnson comments:
Claims that natural selection is a force of stupendous creative
power, which is capable of crafting the immense complexity of biological structures
that living creatures possess in such abundance, are not supported by
experimental evidence or observation. . . Observational evidence (e.g. the
famous peppered moth study) shows mainly cyclical changes in the relative
frequency of characteristics already present in the population. There is
circumstantial evidence pointing to somewhat more impressive changes (e.g.
circumpolar gulls, Hawaiian fruit-fly species), but the empirical evidence
gives no reason for confidence that natural selection has the creative power,
regardless of the amount of time available, to build up complex organs from
scratch or to change one body plan into another. . . The decisive
disconfirmation of neo-Darwinism comes from the fossil record. Even if we
generously grant the assumption that neo-Darwinist macroevolution is capable of
producing basic changes, it does not appear to have done so.[237]
Moreover, as
Ralph O. Muncaster reports: ĹThe technical advances of the latter part of the
twentieth century have allowed scientists to recognize the challenges to the
evolutionary process at the molecular level.ĺ[238]
Climbing
Mount Improbable
According to
Darwinian theory biological systems evolve through the incremental accumulation
of beneficial mutations: Ĺnatural selection acts only by taking advantage of
slight successive variations; she can never take a great a sudden leap, but
must advance by short and sure, though slow stepsĺ.[239] Dawkins
explains why: ĹThe larger the leap through genetic space, the lower the
probability that the resulting change will be viable, let alone an improvement.
[Hence] evolution must in general be a crawl through genetic space, not a
series of leaps.ĺ[240]
He describes this gradual approach to obtaining biological complexity as
ĹClimbing Mount Improbableĺ.[241]
Improbable because, as Steven Vogel writes, the theory stipulates that: ĹNature
in effect must transmute a motorcycle into an automobile while providing
continuous transportation. The need for growth without loss of function can
impose severe geometrical limitations.ĺ[242] As Stephen Jay Gould admitted: ĹOur
inability, even in our imagination, to construct functional intermediaries in
many cases has been a persistent and nagging problem for gradualistic accounts
of evolution.ĺ[243]
Gould also pointed out that: ĹThe fossil record with its abrupt transitions
offers no support for gradual change. . . palaeontologists know that the fossil
record contains precious little in the way of intermediate forms; transitions
between major groups are characteristically abrupt.ĺ[244]
Nevertheless, naturalists, who assume that evolution is true because it
fills the explanatory gap left by the exclusion of design, have been
content to say that even though we have no idea what path organisms took up
Mount Improbable, they must have done so.
For example, Dawkins says that: Ĺhowever daunting the sheer cliffs
that the adaptive mountain first presents, graded ramps can be found the other
side and the peak eventually scaledĺ.[245] How does he know that these Ĺgraded ramps
can be foundĺ in advance of showing what they are; without even looking for
them? Because his justification for this assumption is philosophical rather
than scientific: ĹWithout stirring from our chair, we can see that it must be
soĺ,[246]
explains Dawkins, Ĺbecause nothing except gradual accumulation could, in
principle, do the job. . .ĺ[247]
What job is that? The job of explaining life naturalistically! As Daniel
Dennet admits: ĹThis is a purely theory-driven explanation, argued a-priori
from the assumption that natural selection tells the true story ľ some true
story or other ľ about every curious feature of the biosphere. . . it assumes
that Darwinism is basically on the right track.ĺ[248] Once
again, Dawkinsĺ conclusion rests upon his presupposition that there is no
designer: Ĺthis sort of argument smacks of tautologyĺ, complains Michael
Denton: ĹThe only warrant for believing that functional living systems are. . .
capable of undergoing functional transformation by random mechanisms, is belief
in evolution by the natural selection of purely random changes in the structure
of living things. But this is precisely the question at issue.ĺ[249]
Absence of evidence for precise causal histories of how Mount
Improbable was supposedly conquered by gradualistic means is not conclusive
evidence of an absence of such ascents (although the burden of proof is on the
hypothesis that such events happened); but while design theorists are biased in
favour of naturalistic explanations where they are adequate, the
improbability of the gradualistic hypothesis, together with its pervasive lack
of evidential support, counts against it as an explanation for anything that
looks designed (at least for anyone without a dogmatic commitment to
design-free explanations). As Dembski explains:
Darwinism is committed to a sequence of manageable steps that
gradually transforms A into B. In consequence, there has to be some sequence. .
. where each transition from one step to the next can readily be accounted for
in terms of natural selection and random variation. Thus, for instance, in a Darwinian
explanation of the bacterial flagellum, we know that bacteria lacking a
flagellum (and also lacking any genes coding for a flagellum) had to evolve
into bacteria with a flagellum (and thus possessing a novel genetic complement
for the flagellum). If Darwinism is correct, some step-by-step Darwinian
process had to take us from the former type of bacteria to the latter. So how
did it happen? How could it have happened? Nature somehow filled in the
details, but Darwinists somehow never do.[250]
Dawkins reports Ĺfeeling. . . that provided the difference between neighbouring
intermediates in our series [of forms] is sufficiently small, the necessary
mutations are almost bound to be forthcomingĺ.[251]
However, we know that: ĹThe DNA replication process for higher organisms
includes proofreading to keep error rates at less than one in ten billion
[while] errors that do occur are almost invariably neutral or harmful.ĺ[252]
For example, Darwinists have failed to provide evidence that the eye
could be evolved from scratch though any series of Ĺsufficiently smallĺ
mutations, let alone to provide evidence that any eye historically followed any
such evolutionary pathway: ĹIngenious hypothetical scenarios for the evolution
of complex adaptations are presented to the public virtually as fact, but
sceptics within science derisively call them ôjust-soö stories, because they
can be neither tested experimentally nor supported by fossil histories.ĺ[253]
Biologist Michael Denton notes that: Ĺthrough what intermediate states the
reflecting eye evolved is a mysteryĺ.[254] Indeed, cell biologist Franklin Harold
admits: ĹThere are presently no detailed Darwinian accounts of the evolution of
any biochemical or cellular system, only a variety of wishful speculations.ĺ[255] Even
the just-so stories tend to Ĺartificially isolate a particular component or
organ, such as the eye, from the immensely complex system [i.e. the organism]
in which it is embeddedĺ.[256]
As Johnson complains: ĹThere is no requirement that any of this speculation be
confirmed by either experimental or fossil evidence. For Darwinists, just being
able to imagine the process is sufficient to confirm that something like that
must have happened.ĺ[257]
Dawkins argues that different forms of eye present in nature can be
arranged into a sequence from less to more complex, and that this sequence
shows the viability of a Darwinian explanation of the eye: Ĺmodern analogues of
every step up the ramp can be found, working serviceably in dozens of eyes
dotted independently around the animal kingdomĺ.[258] However,
not only is this sequence of eyes historically hypothetical (showing at best
what could rather than what did happen), but its supposedly simple beginning
actually assumes the existence of Ĺa cell sensitive to light and able to react
in a specific way to visible radiation. . .ĺ[259] That light sensitive cell, far from being
simple, is an example of an Ĺirreducibly complexĺ (IC) system that canĺt have
evolved directly by incremental modifications of a precursor system, because
all of its parts are necessary to its performing its function. As Jakob Wolf
complains: Ĺfar from commanding the resources to explain their emergence,
Darwinian explanation consistently presupposes the existence of irreducibly
complex systemsĺ.[260]
Hence recent discoveries in microbiology mean that the pervasive absence of
evidence for the climbing of ĹMount Improbableĺ has now been transmuted into
evidence of absence:
Climbing Mount Improbable requires taking a slow serpentine route up
the backside of the mountain and avoiding precipices. For irreducibly complex
systems that have numerous diverse parts and that exhibit the minimal level of
complexity needed to retain a minimal level of function, such a gradual ascent
up Mount Improbable is no longer possible.[261]
For some
biological systems (systems exhibiting IC), the task of climbing ĹMount Improbableĺ is to all intents and purposes the task of climbing Mount Impossible. If a biological system exhibits irreducible complexity, then evolution by natural
selection is in serious trouble as an adequate explanatory hypothesisá (and
intelligence, as the only cause known to be capable of originating irreducibly
complexity, moves to the fore as the best explanation of IC systems).
Place Your Bets
Darwin argued that the existence of a single irreducibly complex system
would falsify his evolutionary hypothesis: ĹIf it could be demonstrated that
any complex organ existed which could not possibly have been formed by
numerous, successive modifications, my theory would absolutely break down.ĺ[262] Darwin was betting that no such system would be discovered.
Richard Dawkins places the same bet when he acknowledges that
Darwinĺs remark Ĺis valid and very wise. . . his theory is indeed falsifiable.
. . and he puts his finger on one way in which it might be falsifiedĺ.[263]
However, he asserts that Ĺnot a single case is known to me of a complex organ
that could not have been formed by numerous slight [un-guided] modifications. I
do not believe that such a case will ever be found.ĺ[264]
Nevertheless, he concedes: ĹIf it is ľ itĺll have to be a really complex organ,
and . . . you have to be sophisticated about what you mean by ôslightö ľ I
shall cease to believe in Darwinism.ĺ[265] Like Darwin, Dawkins has a lot riding on
the universal negative bet that nothing in nature is irreducibly complex.
Massimo Pigliucci goes further than Darwin and Dawkins, writing that: Ĺirreducible
complexity is indeed a hallmark of intelligent design.ĺ[266] If
there is irreducible complexity in living organisms, then Pigliucci would
accept this as evidence of intelligent design: Ĺirreducible complexity is
indeed a valid criterion to distinguish between intelligent and nonintelligent
design.ĺ[267]
However, like Darwin and Dawkins, Pigliucci thinks that: Ĺthere is no evidence
so far of irreducible complexity in living organisms.ĺ[268] Note
that the issue here is a dispute about evidence, about the empirically knowable
facts of the matter ľ it is, in that sense, a scientific dispute.
The
Irreducible Behe
ĹBehe . . .
does have a point concerning irreducible complexity. . .ĺ ľ Massimo Pigliucci[269]
The concept of irreducible
complexity is not new. Nor are design arguments based upon the concept. Popular
summaries of William Paley notwithstanding, his argument goes well beyond
analogy, however impressive. Although he didnĺt employ this precise
terminology, Paley pointed out that a watch is, at its core, irreducibly
complex. That is, not only is the watchĺs purpose carried out by the sum of
the watchĺs parts, but that purpose could not be carried out Ĺif its different
parts had been differently shaped from what they are, or placed after any other
manner or in any other order than that in which they are placed . . .ĺ[270] In the contemporary
literature of design, a system performing a given basic function (like a watch
telling the time), is said to be Ĺirreducibly complexĺ if it includes a set of
well-matched, mutually interacting, non-arbitrarily individuated parts such
that each part in the set is indispensable to maintaining the systemĺs basic
function. The set of indispensable parts is the irreducible core of the
system in question.[271]
ĹThe power of the concept of irreducible complexityĺ, writes John A. Smart, Ĺis
that it invalidates the step-by-step process of evolution.ĺ[272] As Behe, Dembski and Meyer
explain: ĹNatural selection can only act on systems that perform functions that
help organisms survive. But ôirreducibly complexö systems have no function at
all unless all the parts in the system are present. Yet without the aid of
natural selection the odds against such systems arising on their own are
prohibitive.ĺ[273]
The existence of an
irreducibly complex system in nature is indicative of design. As atheist Daniel
Dennet, who called such systems Ĺthe You-Couldnĺt-Get-Here-From-There Organ or
Organism,ĺ[274]
explains: ĹIf there are designs that cannot be approached by a gradual,
stepwise redesign process in which each step is at least no worse for the
geneĺs survival chances than its predecessor, then the existence of such a
design in nature would seem to require, at some point in its ancestry, a
helping hand from a foresighful designer . . .ĺ[275] In 1945 British philosopher
A.E. Taylor argued that what made an inference to design irresistible in the
case of a watch was: Ĺthe way in which the various parts of the watch are
co-adapted to produce a unitary result, and a result which cannot be effected
until they are all assembled in a definite way.ĺ[276] Taylor argued that Ĺthe
thorough-going co-adaptation of the parts of organisms to contribute to a
unitary result which will only emerge when the organism is mature may be
ascribed to ôprospective contrivanceö with an even higher degree of
probability.ĺ[277]
University biochemist Michael J. Behe
has put new life into the concept of irreducible complexity by applying the
argument at the bio-molecular level of cellular machinery unknown even in Taylorĺs generation. Beheĺs most notable presentation of irreducible complexity is Darwinĺs
Black Box: the Biochemical Challenge to Evolution (1996), where he defined
his terms as follows:
By irreducibly complex I mean a single system composed of
several well-matched, interacting parts that contribute to basic function,
wherein the removal of any one of the parts causes the system to effectively
cease functioning. An irreducibly complex system cannot be produced directly .
. . by slight, successive modifications of a precursor system, because any
precursor to an irreducibly complex system that is missing a part is by
definition non-functional.[278]
Mike Gene
reports that: ĹBeheĺs concept of irreducible complexity (IC) has found itself
in the peer-reviewed scientific literature. . . and is being taken seriously by
scientists.ĺ[279]
 |
In Darwinĺs Black Box (Simon & Schuster, 1998), Professor Michael J. Behe argues that the cell contains many Ĺirreducibly complexĺ bio-molecular machines, that such machines are unlikely candidates for Darwinian explanation and good candidates for explanation in terms of Ĺintelligent designĺ. |
By
definition, any system that is irreducibly complex
cannot have evolved directly up any Ĺgraded rampĺ round the back of ĹMount Improbableĺ. Ruling out direct, incremental evolution does not exclude what Darwin called Ĺa sudden leapĺ,[280] but as Dawkins notes: ĹThe larger the leap through genetic space, the lower the
probability that the resulting change will be viable, let alone an
improvement.ĺ[281]
Behe observes:
Even if a system is irreducibly complex (and thus cannot have been
produced directly). . . one can not definitely rule out the possibility of an
indirect, circuitous route. As the complexity of an interacting system
increases, though, the likelihood of such an indirect route drops
precipitously. And as the number of unexplained, irreducibly complex biological
systems increases, our confidence that Darwinĺs criterion of failure has been
met skyrockets. . .[282]
Sceptics of
Beheĺs hypothesis ideally need to demonstrate the existence of indirect,
Ĺcircuitousĺ, graded evolutionary pathways with the following features: they
achieve the gradual assembly of all and only the components required by a given
IC system (in the correct, functionally specific arrangement) through a
sufficiently probable series of evolutionary steps, wherein each and every step
a) has a function, b) either increases the fitness of the preceding functional
step or results in a system with a different advantageous function, and where
c) all of these functional steps are (by definition) different to the function
exhibited by the IC end-product being explained. But as Dembski observes:
The fact is that for irreducibly complex biochemical systems, no
indirect Darwinian pathways are known. . . Whatĺs needed is a seamless
Darwinian account thatĺs both detailed and testable of how subsystems
undergoing co-evolution could gradually transform into an irreducibly complex
system. No such accounts are available or forthcoming. . . The absence here is pervasive
and systemic. . .[283]
Critics charge Beheĺs argument with failing to acknowledge that
Ĺabsence of evidence is not evidence of absenceĺ, but this truism has its
limitations. Failing to find car oneĺs keys in the house doesnĺt indicate they
donĺt exist: after all, they might be in the car and you know you own keys to
be discovered. On the other hand, suppose one wasnĺt sure that keys existed to
be discovered, and had conducted a thorough search anyway, and had come up
empty handed. The latter analogy is closer to the scientific reality of
explaining IC systems. As Dembski observes:
If after repeated attempts looking in all the most promising places
you donĺt find what you expect to find and if you never had any evidence that
the thing you were looking for existed in the first place, then you have reason
to think that the thing you are looking for doesnĺt exist at all. . . the
absence of scientific evidence here is as complete as it is for leprechauns.[284]
Moreover, as
Robert C. Koons argues, the burden of proof here: Ĺis on Darwin and his
defenders to demonstrate that it is really possible for at least some of the
complex organs we find in nature to be formed in this way: that is, by some
specific, fully articulated series of slight modifications.ĺ[285] Darwin was right to note that demonstrating the existence of a single biological system
that could not possibly have been formed by numerous, successive modifications,
would cause his theory to Ĺabsolutely break down.ĺ[286]
However, he was wrong to set the evidential bar quite so high (demonstrating
the existence of a system that is highly unlikely to have been formed by
numerous, successive - and unguided - modifications would cause a sufficiently
catastrophic break down in his theory); and he was doubly wrong to suggest that
the evolutionary hypothesis should be extended the presumption of truth until
such a demonstration could be made:
the inference from complex, interdependent functionality to
intelligent agency is the natural, default position. Darwinian biologists and
their pupils overlook this fact at their own cognitive peril. . . The bare
possibility that a non-teleological explanation of apparent design might exist
is not by itself sufficient to warrant real doubt about the reality of design,
any more than the bare possibility that you and I are brains in a Matrix-like
vat is sufficient to warrant scepticism about the deliverances of our five
senses.[287]
As Koons
writes elsewhere:
The burden is on the skeptic to provide good rational or empirical
grounds for believing that the false appearance of design is not unlikely under
actual conditions. Design sceptics, in common with other sceptics from
antiquity to the modern day, attempt an illegitimate shift in the burden of
proof. The skeptic attempts to rebut a successful design inference by merely raising
the possibility that the appearance of design may be illusory, challenging
the defender of the inference to prove a negative ľ to prove that the sceptical
scenarios could not have happened. The appropriate response to such
sceptical challenges is to place the burden of proof where it belongs: the
skeptic must provide substantial and specific grounds for doubting the
soundness of the design inference in the particular case in question.[288]
Nevertheless, Behe graciously shoulders the burden of proof that
should rightfully apply to the Darwinian, and argues that at the biomolecular
level of life (which was an unknown ĹBlack Boxĺ in Darwinĺs day) there are
several irreducibly complex molecular systems that are highly unlikely
to have been formed by numerous, successive (and unguided) modifications:
Ĺincluding aspects of protein transport, blood clotting, closed circular DNA,
electron transport, the bacterial flagellum, telomeres, photosynthesis,
transcription regulation, and much moreĺ.[289]
Given that such systems are resistant to naturalistic explanations,
and given our everyday experience that intelligent agents regularly produce
irreducibly complex systems, Behe goes on to argue that the best explanation of
such molecular machines is intelligent design:
When is it reasonable to conclude, in the absence of firsthand
knowledge or eyewitness accounts, that something has been designed? For
discrete physical systems ľ if there is not a gradual route to their production
ľ design is evident when a number of separate, interacting components are
ordered in such a way as to accomplish a function beyond the individual
components. The greater the specificity of the interacting components required
to produce the function, the greater is our confidence in the conclusion of
design.[290]
Consider just
one example of an IC system discussed by Behe:
The Fantastic
Flagellum
[cf. E. Coli
swimming with flagella @
www.mtmi.vu.lt/pfk/funkc_dariniai/nanostructures/bacteria.htm]
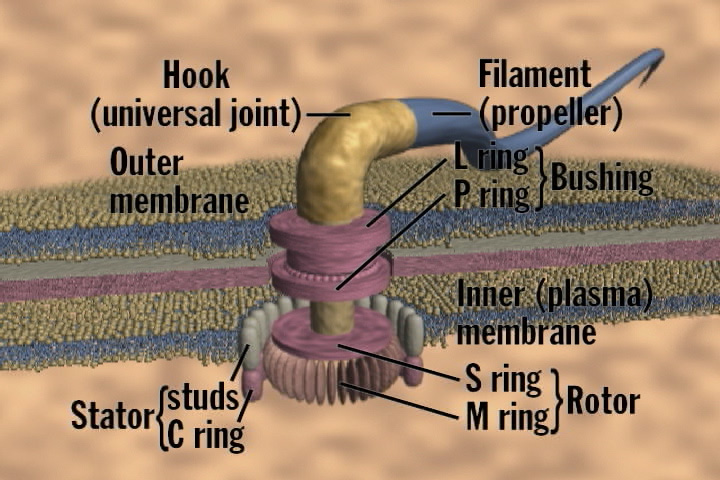 In 1973 the
startling, unexpected discovery was made that some bacteria swim using an
outboard motor, spinning their Ĺflagellaĺ at up to 100,000 rpm like a screw
propeller: ĹThe rotary motor, with a diameter of only 30 to 40 nm [nanometers],
drives the rotation of The Bacterial Flagellum the
flagellum at around 300 Hz, at a power level of 10-16 W with energy
conversion efficiency close to 100%.ĺ[291]
In 1973 the
startling, unexpected discovery was made that some bacteria swim using an
outboard motor, spinning their Ĺflagellaĺ at up to 100,000 rpm like a screw
propeller: ĹThe rotary motor, with a diameter of only 30 to 40 nm [nanometers],
drives the rotation of The Bacterial Flagellum the
flagellum at around 300 Hz, at a power level of 10-16 W with energy
conversion efficiency close to 100%.ĺ[291]
(Labelled Illustration of bacterial flagellum from ARN Molecular Museum)
The
bacterial flagellum is irreducibly complex: ĹThe flagellum includes an acid
powered rotary engine, a stator, O-rings, bushings and a drive shaft. The
intricate machinery of this molecular motor requires approximately [forty]
proteins. Yet the absence of any one of these proteins results in the complete
loss of motor function.ĺ[292]
One can see that a rotary motor without a propeller, or a drive shaft, or a
motor, just wonĺt work: ĹBecause the bacterial flagellum is necessarily
composed of at least three parts ľ a paddle, a rotor, and a motor ľ it is
irreducibly complex.ĺ[293]
Robert Macnab comments:
one can only marvel at the intricacy, in a simple
bacterium, of the total motor and sensory system which has been the subject of
this review and remark that our concept of evolution by selective advantage
must surely be an oversimplification. What advantage could derive, for example,
from a Ĺpreflagellumĺ (meaning a subset of its components), and yet what is the
probability of Ĺsimultaneousĺ development of the organelle at a level where it
becomes advantageous?[294]
Experiments
have confirmed that eliminating any of the proteins that form the flagellum
results in a non-functioning machine. That is, the flagellum is irreducibly
complex, a fact that rules out explanation by direct evolutionary
pathways and leaves the Darwinist to fall back on far chancier indirect
explanations.
How the Flagellum Evolved? A Just-So story[295]
ĹI have learned
from my own embarrassing experience how easy it is to concoct remarkably
persuasive Darwinian explanations that evaporate on closer inspection.ĺ ľ
Daniel Dennet[296]
Christian biologist
Ken Miller has argued for an indirect explanation in the case of the flagellum
by pointing to the existence of the type III secretory system (TTSS),[297] which
is coded for by about ten genes, each of which is structurally similar
(homologous) to genes in the bacterial flagellum. Miller sees the TTSS as a
functional evolutionary precursor of the flagellum capable of being selected
for on its own functional merits and then augmented to produce the flagellum.
However, as Dembski points out: ĹAt best the TTSS represents one possible step
in the indirect Darwinian evolution of the bacterial flagellum. Whatĺs needed
is a complete evolutionary path and not merely a possible oasis along the way.
To claim otherwise is like saying we can travel by foot from Los Angeles to Tokyo because weĺve discovered the Hawaiian Islands.ĺ[298] Two final points nail shut the coffin of
Millerĺs TTSS scenario. The first is that ĹThe type III system itself is [IC],
perhaps with ten IC components.ĺ[299]
The second is that: ĹThe best current molecular evidence. . . points to the
TTSS evolving from the flagellum and not vice versa.ĺ[300]
H. Allen Orr (a critic of Behe) argues:
First, it will do no good to suggest that all the required parts of
some biochemical pathway popped up simultaneously by mutation. Although this
Ĺsolutionĺ yields a functioning system in one fell swoop, itĺs so hopelessly
unlikely that no Darwinian takes it seriously. . . Second, we might think that
some of the parts of an irreducibly complex system evolved step by step for other
purposes and were then recruited wholesale to a new function. But this is also
unlikely.[301]
Nevertheless,
the current favourite among indirect explanations being advanced for IC systems
is Orrĺs second option of the wholesale co-option of parts evolved for other
functions, an option R. H. Thornhill and D. W. Ussery call Ĺadoption from a
different functionĺ.[302]
While the TTSS, for example, contains around ten proteins that are homologous
to proteins in the flagellum, the flagellum has another thirty or so proteins,
which are unique to it. As Scott Minnich, Professor of Biology at the University of Idaho, and an expert on the flagellum, says in response to the co-option
hypothesis:
With a bacterial flagellum, youĺre talking about a machine thatĺs
got forty structural parts. Yes, we find ten of them are involved in another
molecular machine, but the other thirty are unique. So where are you going to
borrow them from? Eventually youĺre going to have to account for the function
of every single part as if originally having some other purpose. I mean you can
only follow the argument so far, until you run into the problem that youĺre
borrowing from nothing. . .ĺ[303]
Stripping away a
third of the improbability Dembski calculates for the spontaneous formation of
the flagellum (10-1170) doesnĺt appreciably improve matters for a
naturalistic explanation: ĹApplied to those remaining two-thirds of flagellar
proteins, my calculation yields something like 10-780, which also
falls well below my universal probability bound.ĺ[304] As Mike Gene warns: ĹThe
brilliance of Darwin was to minimize the role of chance in apparent design. But
once we turn to the co-option explanation, we leave this explanatory appeal
behind, as chance reasserts itself into a place of prominence.ĺ[305]
Taking what Joshua A. Smart calls Ĺa Concessionary approachĺ[306], Dembski supposes, purely for
the sake of argument, that we discover several molecular systems Ĺlike the TTSS
that jointly took into account all the flagellar proteinsĺ.[307] Those proteins would be
Ĺsimilar but, in all likelihood, not identical to the flagellar proteins
(strict identity would itself be vastly improbable)ĺ.[308] Such a hypothetical
situation, designed to maximize the chances of an indirect explanation by
co-option, Ĺraises the question how those several molecular machines can come
together so that proteins from one molecular machine adopt proteins from
another molecular machine to form an integrated functional system like the
flagellumĺ.[309]
As Minnich says: Ĺeven if you concede that you have all the parts necessary to
build one of these machines, thatĺs only part of the problem. Maybe even more
complex is the assembly instructions.ĺ[310]
Dr John Bracht, managing editor of the journal Progress in Complexity,
Information and Design,[311]
explains:
biological functionality is turning out to be much more highly
specified and precise than we had originally envisioned. . . biology is really
a science of engineering, where the constraints for bio-functionality are
extreme ľ to the point that nearly every molecular interaction is remarkably
precise and tightly controlled. Molecular biology is much like a jigsaw puzzle
where each piece must be specifically shaped to fit with the other pieces
around it. . .[312]
Applying these
observations to the proposed construction of the flagellum by co-option, Bracht
writes:
The problem is that the proteins which are to become the flagellum
are coming from systems that are distinctly non-flagellar in nature. . . and
being co-modified from their original molecular interactions into an entirely
new set of molecular interactions. Old interfaces and binding sites must be
removed and new ones must be created. But given the sheer number of flagellar
proteins that must co-evolve. . . the Darwinian explanation is [very unlikely
and therefore] really no different from appealing to a miracle.[313]
Dembski adds
the observation that: Ĺthe only evidence we have of successful co-option comes
from engineering and confirms that intelligence is indispensable in explaining
complex structures like. . . the bacterial flagellum,ĺ[314] and he
concludes: ĹWe can do the probabilistic analysis at the level of individual
proteins. . . Or we can do it at higher levels of organization like functional
subsystems [like TTSS]. . . But all such probabilistic analyses still point up
vast improbabilities.ĺ[315]
The
Origination Probability and the Origination Inequality
In
ĹIrreducible Complexity Revisitedĺ, a paper reviewing the status of Beheĺs
concept of irreducible complexity eight years on from the publication of Darwinĺs
Black Box, William A. Dembski suggests that Ĺthe problem he has raised is,
if anything, still more vexing for Darwinism that when he first raised it.ĺ
Dembski observes that: Ĺeven when the most generous allowance of legitimate
advantages, the probabilities computed for the Darwinian mechanism to evolve
irreducibly complex biochemical systems like the bacterial flagellum always end
up being exceedingly small.ĺ[316]
He goes on to explain why this is the case, arguing that the reason these
probabilities always end up being so small: Ĺis the difficulty of coordinating
successive evolutionary changes apart from teleology. . . In attempting to
coordinate the successive evolutionary changes needed to bring about
irreducibly complex biomechanical machines, the Darwinian mechanism therefore
encounters a number of daunting probabilistic hurdles.ĺ[317] These
hurdles include:
- Availability. Are
all the parts needed to evolve an IC system even available?
- Synchronization.
Are all these parts available at the right time so that they can be
incorporated into the evolving structure?
- Localization.
Even with all the right parts available at the right time, can they all be made
available at the Ĺconstruction siteĺ of the evolving system?
- Interfering Cross-Reactions. Given that all the right parts can all be brought together at the
right time and in the right place, how can the wrong parts that would otherwise Ĺgum upĺ the works be excluded from the Ĺconstruction siteĺ?
-
Interface Compatibility. Will all the parts being recruited for inclusion in an evolving
system be compatible in the sense of interfacing tightly so that, once suitably
positioned, they work together to form a functioning system?
- Order of Assembly.
Even if all and only the right parts reach the right place at the right time,
and even if they have full interface compatibility, will they be assembled in
the right order to form a functioning system?
Dembski uses
an analogy to explain what is at stake in overcoming these probabilistic
hurdles. A building contractor building a house needs to overcome each of the
hurdles listed above. A building contractor doesnĺt find this task
insurmountable. Thatĺs because, as an intelligent agent, they have the know-how
to think ahead and to coordinate all the tasks needed to clear these hurdles with
reference to achieving the goal of building a house:
But the Darwinian mechanism of random variation and natural
selection has none of this know-how. All it knows is how to randomly modify
things and then preserve those random modifications that happen to be useful at
the moment. The Darwinian mechanism is an instant gratification machine. If the
Darwinian mechanism were a building contractor, it might put up a wall because
of its immediate benefit in keeping out intruders from the construction site even
though by building the wall now, no foundation could be laid later and, in
consequence, no usable house could ever be built at all. Thatĺs how the
Darwinian mechanism works, and thatĺs why it is so limited.[318]
Is the limited
know-how of the Darwinian mechanism adequate for the task of coordinating the
biochemical events needed to clear all these hurdles? ĹTo answer yes to this
question,ĺ says Dembski, Ĺis to attribute creative powers to the Darwinian
mechanism that are implausible in the extreme.ĺ[319]
Considering each of the hurdles in turn gives a good idea of just how
implausible:
(1) Availability
To clear this hurdle, the Darwinian mechanism needs to be able to
form novel proteins from scratch. . . the Darwinian mechanism is capable of
tinkering with existing proteins or recruiting them wholesale for new uses. But
there is no evidence that it can produce complex specified proteins from
scratch. . . recent work on the extreme functional sensitivity of proteins
provides strong evidence that certain classes of proteins are in principle
un-evolveable by gradual means (and thus a fortiori by the Darwinian
mechanism) because small perturbations of these proteins destroy all
conceivable biological function (and not merely existing biological function).
Thus, it is highly implausible that the Darwinian mechanism can generate the
novel proteins. . . required in the evolution of the bacterial flagellum.[320]
(2) Synchronization
Some hurdles are easier for the Darwinian mechanism to clear than
others, and this is perhaps one of them. . . The only hitch could be that an
item that hitherto has served a biologically useful function and is needed in
the future evolution of some irreducibly complex system loses its functional
advantage somewhere in the middle of the evolutionary process and thus falls
into disuse. If that happens, natural selection will tend to eliminate that
item, thereby rendering it unavailable.[321]
(3) Localization
The localization hurdle. . . seems considerably more
difficult for the Darwinian mechanism to clear. . . items originally assigned
to certain systems need to be reassigned and recruited for use in a newly
emerging system. . . But how likely is it that these items break free and get
positioned at the construction site of an existing system, thereby transforming
it into a newly emerging system with a novel or enhanced function? Our best
evidence suggests that this repositioning of items previously assigned to
different systems is improbable and becomes increasingly improbable as more
items need to be repositioned simultaneously at the same location.[322]
(4) Interfering
Cross-Reactions
by welcoming items that could help in the evolution of the bacterial
flagellum, the construction site would also welcome items that could hinder its
evolution. It follows that to the degree that the localization hurdle is easy
to clear, to that degree the interfering cross-reaction hurdle is difficult to
clear, and vice versa.[323]
(Hurdles 3 & 4 constitute a ĹCatch 22ĺ situation for evolution.)
(5) Interface
Compatibility
With the interface-compatibility hurdle, we come to the
gravest difficulty confronting the Darwinian mechanism. . . if these items
[used in constructing bio-molecular systems] were built according to common
standards or conventions, there might be some reason to think that they could
work together effectively. But natural selection is incapable of instituting
such standards or conventions. . . In fact, common standards and conventions
that facilitate the interface compatibility of distinct functional systems points
not just to the design of the systems but also to a common design responsible
for the common standards and conventions. . . As an instant gratification
mechanism [the Darwinian mechanism] has no stake in ensuring that such
structures also adhere to standards and conventions that will allow them to
interface effectively with other structures down the line. . . there is no
evidence, whether theoretical or experimental, that the Darwinian mechanism can
clear the interface compatibility hurdle.[324]
(6) Order
of Assembly
For a Darwinian mechanism to clear the order-of-assembly hurdle
is also a stretch. . . in the evolution of [IC] systems. . . we can expect the
order of assembly of parts to undergo substantial permutations. . . How, then,
does the order of assembly undergo the right permutations? For most biological
systems, the order of assembly is entrenched and does not permit substantial
deviations. The burden of evidence is therefore on the Darwinist to show that
for an evolving system, the Darwinian mechanism coordinates not only the
emergence of the right parts but also their assembly in the right order.
Darwinists have done nothing like this.[325]
Dembski notes
that it is possible to assess objectively and quantitatively the challenge that
these hurdles pose to the Darwinian mechanism. Associated with each hurdle is a
probability P [i.e. Pavail, Psynch, Plocal, Pi-c-r,
Pi-f-c, Po-o-a]. Each of these probabilities is conditional on
the proceeding ones. For example, the synchronization probability (Psynch)
assesses the probability of synchronization on condition that the needed
parts are available (Pavail). Consequently, the probability of an IC
system arising by Darwinian means cannot exceed the following product:
Pavail x Psynch x Plocal x
Pi-c-r x Pi-f-c x Po-o-a
If we define Porigin
as the probability of an IC system originating by Darwinian means (the origination
probability), Ĺthen the following inequality holds (the origination
inequality)ĺ[326]:
Porigin < Pavail
x Psynch x Plocal x Pi-c-r x Pi-f-c x Po-o-a
(That is, the
origination probability is equal to or less than the product.) This inequality
tells us that: Ĺeven if one of the probabilities to the right of the inequality
sign is small, then the origination probability must itself be small (indeed, no
bigger than any of the probabilities on the right).ĺ[327] We
neednĺt calculate all the probabilities to the right of the inequality sign to
ensure that Porigin is small:
It is enough to have reliable upper bounds on these probabilities.
If any one of these upper bounds is small, then so is the associated
probability and so is the origination probability. And it the origination
probability is small, then the irreducibly complex system in question is both
highly improbably and specified (all these irreducibly complex systems are
specified in virtue of their biological function). It follows that if the
origination probability is small, then the system in question exhibits
specified complexity; and since specified complexity is a reliable empirical
marker of actual design, it follows that the system itself is designed.[328]
Hence
irreducible complexity is revealed to be a concrete example of specified
complexity:
The irreducibly complex systems Behe considers require numerous
components specifically adapted to each other and each necessary for function.
On any formal complexity-theoretic analysis, they are complex in the sense
required by the complexity-specification criterion. Moreover, in virtue of
their function, these systems embody patterns independent of the actual living
systems. Hence these systems are also specified in the sense required by the
complexity-specification criterion.[329]
Dembski
summarises the argument for design from IC systems:
irreducible complexity renders biological structures provably inaccessible
to direct Darwinian pathways. . . the failure of evolutionary biology to
discover indirect Darwinian pathways is pervasive and systemic and therefore
reason to doubt. . . that indirect Darwinian pathways are the answer to
irreducible complexity. . . itĺs not just that certain biological systems are
so complex that we canĺt imagine how they evolved by Darwinian pathways.
Rather, we can show conclusively that direct Darwinian pathways are causally
inadequate to bring them about and that indirect Darwinian pathways [which seem
unlikely on both theoretical and experimental grounds and] are utterly without
empirical support in bringing them about [and so fail to meet the proper burden
of proof]. Conversely, we do know what has the causal power to produce irreducible
complexity ľ intelligent design.[330]
Intelligent
design looks like the best ľ most causally adequate ľ explanation for IC
systems such as the flagellum, and Darwinists appears to be betting in the face
of some long odds: ĹLike compulsive gamblers who are constantly hoping that
some really big score will cancel their debts, evolutionary biologists live on
promissory notes that show no sign of being redeemable. . .ĺ[331] If
biologists can meet the burden of proof by discovering or constructing
detailed, testable, indirect Darwinian pathways of sufficiently high
probability that account for the emergence of irreducibly and minimally complex
biological systems like the bacterial flagellum, Ĺthen more power to themĺ,[332] says
Dembski (the assertion that a given system is IC is scientifically
falsifiable): ĹBut until that happens, evolutionary biologists who claim that
natural selection accounts for the emergence of the bacterial flagellum are
worthy of no more credence than compulsive gamblers who are forever promising
to settle their accounts.ĺ[333]
Maximally
IC Systems
IC systems
like the bacterial flagellum are in principle inaccessible to direct
Darwinian pathways, and evolutionary biologists therefore pin their hopes on
the possibility that sufficiently probable indirect Darwinian pathways
might one day be found to account for them. (To the best of our knowledge the
odds do not favour such explanations, and hence the best explanation of such
systems is intelligent design.) However, recent research points to the
existence of complex biological systems that are not only inaccessible in
principle to direct Darwinian pathways (making them Ĺminimallyĺ IC), but
inaccessible in principle to indirect pathways as well (making them Ĺstronglyĺ
or Ĺmaximallyĺ IC). As Dembski explains:
there is now mounting evidence of biological systems for which any
slight modification does not merely destroy the systemĺs existing function but
also destroys the possibility of any function of the system whatsoever.
(Consult, for instance, the research on extreme functional sensitivity of
various enzymes and on irreducibly complex metabolic pathways of enzymes for
which each enzyme needs to attain a certain catalytic threshold before it or
its associated pathway can serve any biological function at all.) For such
systems, neither direct nor indirect Darwinian pathways could account for them.[334]
Beheĺs remark
that Ĺif a system is irreducibly complex (and thus cannot have been produced
directly). . . one can not definitely rule out the possibility of an indirect,
circuitous routeĺ[335]
applies to Ĺminimally irreducibly complexĺ systems, but not to Ĺmaximally
irreducibly complexĺ systems. When it comes to a maximally IC system (and that
includes otherwise minimally IC systems that depend upon any maximally IC
systems), anyone suggesting that Darwinian mechanisms alone can offer a
sufficient explanation would not only be betting against the odds, but against
an in principle argument to the contrary. As Dembski observes: Ĺno
Darwinian account could in principle be given for the emergence of such
systemsĺ.[336]
Irreducible
Complexity & Darwinĺs Extrapolation
The existence
of irreducibly complex bio-molecular systems, systems that are highly
unlikely to have been formed by numerous, successive, unguided modifications,
causes Darwinĺs extrapolation from micro to macro-evolution to Ĺabsolutely
break downĺ[337]
because they demonstrate the falsity of his assumption that there are no
limiting factors preventing the mechanism of natural selection from explaining
the full panoply of biological diversity as well as observed micro-evolutionary
adjustments. Explaining IC systems by reference intelligent design, on the
other hand, constitutes a causally adequate explanation that is both
intuitively obvious (having the presumption of truth on its side) and is
supported by design-detection criteria accepted by theists (e.g. Dembski) and
atheists (e.g. Dawkins, Pigliucci) alike.
Conclusion:
Prospects for Darwinism & Design
Dawkins
dismisses ID theorists as: Ĺa well-organised and well-financed group of
nuttersĺ.[338]
But this sort of bluster (a debating tactic called ĹPoisoning the wellĺ) should
not intimidate us. Dawkinsĺ bark is worse than his bite (just as his
philosophical skills are worse than his rhetorical skills). The fact of the
matter is that Ĺthe epistemic status of evolutionary biology is, to say the
least, a highly controversial topicĺ[339]
and Ĺa growing number of highly educated scientists. . . are becoming
increasingly sceptical of evolutionary theory.ĺ[340] Thomas Dubay reports: ĹThe
movement in the mainline scientific community to see design in the visible
universe, and most particularly in cellular biology, is growing in strength,
while its opponents have nothing but feeble and unpersuasive responses.ĺ[341] Professor Frederick Crews
grudgingly acknowledges that: ĹIntelligent design is thriving in cultural
circles where illogic and self-indulgence are usually condemned.ĺ[342]
Dembski counters Dawkinsĺ ad hominem attack: Ĺone can be
reasonably well-adjusted, remarkably well-educated (as many design theorists
are), and still think Darwinism is a failed scientific paradigm.ĺ[343] Dembski himself has a B.A in
psychology, an M.S in statistics, a PhD in the philosophy of science and a PhD
in mathematics (all from the University of Chicago), as well as a Master of
Divinity from Princeton Theological Seminary; and he is associate research
professor in the conceptual foundations of science at Baylor University.
According to Dembski:
The following problems have proven utterly intractable not only for
the mutation-selection mechanism but also for any other undirected natural
process proposed to date: the origin of life, the origin of the genetic code,
the origin of multicellular life, the origin of sexuality[344], the scarcity of transitional
forms in the fossil record[345],
the biological big bang that occurred in the Cambrian era[346], the development of complex
organ systems and the development of irreducibly complex molecular machines.
These are just a few of the more serious difficulties that confront every
theory of evolution that posits only undirected natural processes. It is thus
sheer arrogance for Darwinists like Richard Dawkins. . . to charge design
theorists with being ignorant or stupid or wicked or insane for denying the
all-sufficiency of undirected natural processes in biology. . .[347]
Dembski is alluding to a book review by Dawkins
in which he affirms: ĹIt is absolutely safe to say that if you meet somebody
who claims not to believe in evolution, that person is ignorant, stupid or
insane (or wicked, but Iĺd rather not consider that).ĺ[348] Dawkins distinguishes between natural
selection (Ĺthe mutation-selection mechanismĺ) as Darwinĺs proposed engine of
evolution (about which he admits: ĹIt is still (just)
possible for a biologist to doubt its importance, and a few claim to.ĺ[349]) on the one hand, and Ĺthe fact of evolution itself, a fact that is proved utterly beyond
reasonable doubtĺ,[350]
on the other. Hence, by Ĺthe fact of evolution itselfĺ Dawkins appears to mean
the multi-faceted belief that life arose millions of years ago due only to
(undirected) natural processes and then diversified from a common ancestor over
millions of years merely by the operation of (undirected) natural processes.
Elsewhere Dawkins circumspectly writes:
We can now assert with confidence that the theory that the Earth
moves round the Sun not only is right for our time but will be right in all
future times even if flat-Earthism happens to become revived and universally
accepted in some new dark age of human history. We cannot quite say that
Darwinism is in the same unassailable class. Respectable opposition to it can
still be mounted, and it can be argued that the current high standing of
Darwinism in educated minds may not last through all future generations. . . we
must acknowledge the possibility that new facts may come to light which will
force our successors of the twenty-first century to abandon Darwinism or modify
it beyond recognition.[351]
What Dawkins
means by ĹDarwinismĺ here probably co-insides with the theory of natural selection
(which he brackets from Ĺthe fact of evolution itselfĺ); but Dawkins affirms
his faith in the survival of what he calls Core Darwinism: Ĺthe minimal
theory that evolution is guided in adaptively non-random directions by the
non-random survival of small random hereditary chances.ĺ[352] And although Dawkins is
prepared to concede that evolutionary theory might evolve in the future, he
certainly isnĺt prepared to admit any hint of design or purpose within biology:
The feature of living matter that most demands explanation is that
it is almost unimaginably complicated in directions that convey a powerful
illusion [Ĺappearanceĺ would be a less pejorative term] of deliberate design. .
.á Adaptations, especially complex adaptations, awake such a powerful hunger
that they have traditionally provided one of the main motivations for belief in
a supernatural Creator. The problem of adaptation, therefore, really was
a big problem, a problem worthy of the big solution that Darwin provided. . .[353]
Note that the
appearance of deliberate design is assumed to be Ĺa powerful illusionĺ,
and that this presumed Ĺillusion of deliberate designĺ is considered to be a
Ĺbig problemĺ precisely because it (indirectly) supports belief in Ĺa
supernatural Creatorĺ!á For Dawkins the atheist, the fact of evolution itself
must remain staunchly naturalistic, whatever Ĺnew facts may come to lightĺ![354] Dawkinsĺ conclusion is driven
by his philosophy rather than by scientific evidence, a situation that leads
him to cast aspersions on the intellectual or moral character of anyone with
the timidity to call into question Ĺthe fact of evolution itselfĺ (something
that can be done by doubting any of its several facets).[355] Surely we should agree with
W.R. Thompsonĺs introduction to the centenary edition of Darwinĺs Origin,
which castigated Ĺscientific men [who] rally to the defence of a doctrine they
are unable to. . . demonstrate with scientific rigor, attempting to maintain
its credit with the public by the suppression of criticism and the elimination
of difficulties, [a situation that] is abnormal and undesirable in science.ĺ[356]
Increasingly
Common Dissent
The truth of the
matter is that Ĺmore and more biologists, biochemists, and other researchers
have raised serious objections to evolutionary theory in recent years.ĺ[357]
 |
William A. Dembski has edited a number of books bringing together intellectuals from a number of disciplines who dissent from Darwinism. Uncommon Dissent: Intellectuals Who Find Darwinism Unconvincing (ISI Books, 2004) includes contributions from philosophers Robert C. Koons and J. Budziszewski, lawyers Phillip E. Johnson and Edward Sisson, mathematician and philosopher Dr. David Berlinski, biochemist Michael J. Behe, biologist Michael J. Denton, biophysicist Cornelius G. Hunter, Professor of Mathematical Physics Frank J. Tipler, and Professor of the Faculty of Sciences at the University of Paris (and member of the French Academy of Sciences), Marcel-Paul Schutzenberger, and others. |
 |
The July/August 1999 edition of Touchstone Magazine was a special issue on intelligent design theory. It contained a collection of well-written, readable articles by leaders of the movement including: Phillip E. Johnson, Michael J. Behe, Paul Nelson, William A. Dembski, Jonathan Wells, Walter L. Bradley, Stephen C. Meyer, Jay Wesley Richards, Nancy Pearcey, and John Wiester. The magazine issue sold so well that the publishers decided to produce an expanded book edition. The book, co-edited by William A. Dembski and James M. Kushiner, includes all the main articles from the magazine together with a new introduction by Dembski and a new closing article by Bruce L. Gordon on the scientific status and future of design-theoretic explanations. |
 |
Stemming from the 1996 Mere Creation conference at Biola University, this Dembski edited volume includes contributions from philosophers William Lane Craig, Stephen C. Meyer, J.P. Moreland, Paul Nelson, Del Ratzsch and John Mark Reynolds, mathematician and philosopher David Berlinski, Professor of law Phillip E. Johnson, physicist Robert Kaita, engineer Walter L. Bradley, physical anthropologist Sigrid Harwig-Scherer, astronomer Hugh Ross, physical chemist Henry F. Schaeffer III, biochemist Michael J. Behe, biologist Siegfried Scherer, and others. |
Gordon Graham, a
Professor of Philosophy at the University of Aberdeen, says he is Ĺequally
unsympathetic to creationism and to any rabidly anti-religious scientism.ĺ[358] According to Graham: Ĺthe
intellectual difficulties facing neo-Darwinism are formidable. . .ĺ[359] He quotes David Stove, whom
he calls Ĺa respected philosopher writing in a professional journalĺ,[360] who remarks:
Most educated people nowadays. . . think of themselves as
Darwinians. If they do, however, it can only be from ignorance: from not
knowing enough about what Darwinism says. For Darwinism says many things,
especially about our species, which are too obviously false to be believed. . .
at least by any educated person who retains any capacity at all for critical
thought on the subject of Darwinism.[361]
ĹSuffice it to
sayĺ, concludes Graham, Ĺthat there are dissenting voices, and dissenting
voices from within biology itself no less than from anthropology, philosophy,
psychology and sociology.ĺ[362]
According to one of the dissenting voices from within philosophy,
Dr. Norman L. Geisler:
it is unreasonable to believe that intelligent life was caused by
nonintelligent natural forces. . . the odds of life beginning by chance are. .
. for all practical purposes, zero. . . it takes more faith to believe in
evolution than it does to believe in a supernatural creator.[363]
Philosopher
William Lane Craig concludes that:
in the absence of a methodological commitment to naturalism, there
really does not appear to be compelling evidence for the neo-Darwinian theory.
On the contrary, there seems to be pretty persuasive evidence that the
neo-Darwinian account cannot be the full story.[364]
Analysing the
theory of evolution without naturalistic assumptions, Alvin Plantinga, one of
todayĺs leading philosophers, concludes: Ĺthat it happened is doubtful;
that it is certain, however, is ridiculous.ĺ[365]
Taken in any substantive sense, Richard Dawkinsĺ assertion that Ĺno
qualified scientist doubts that evolution is a factĺ[366] is simply incorrect. For
example:
- Paul Chien
(PhD in biology from the university of California, Irvine) is a professor
(and ex-chair) in the Department of Biology at the University of San Francisco. Chien is sceptical about evolution (cf. ĹThe Cambrian Explosion: Biologyĺs
Big Bangĺ, co-authored with historian and philosopher of science Stephen
C. Meyer, palaeontologist Marcus Ross and philosopher of biology Paul
Nelson, in the peer reviewed volume by John Angus Campbell (ed.) Darwinism,
Design, And Public Education, Michigan State University Press, 2003)
and he recently translated Phillip E. Johnsonĺs Darwin on Trial
into Chinese.
- Jonathan Wells
(PhD, Yale; PhD Berkeley) is a post-doctoral biologist in the Department
of Molecular and Cell Biology at the University of California at Berkley. According to Wells: ĹThe Darwinian paradigm is in serious trouble, of the kind
that matters most in science: it doesnĺt fit the evidence.ĺ[367]
 |
In Icons Of Evolution: Science or Myth? (Regnery Publishing, 2002) biologist Jonathan Wells investigates the 10 best known evidences for macro-evolution (e.g. peppered moths, mutations in fruit flies, Haekelĺs embryos, Stanley Millerĺs origin of life experiment, Lucy, homology in limbs, Darwin's finches, horse evolution, etc), and argues that in each case the Ĺevidenceĺ is either fraudulent or falsely interpreted. |
- Chemist Dr.
Cliff Marsh writes:
the factual basis for evolution is, at best, seriously lacking.
Knowledge is growing in all fields of science, but, paradoxically, there is
less and less solid evidence for evolutionary mechanisms. . . The evidence for
evolution being the process by which life originated and developed isnĺt that
convincing. The latest discoveries in biology, chemistry and other areas have
raised some serious questions.[368]
- Dr. Michael J.
Denton, a molecular biologist, writes that he is: Ĺsceptical that major
evolutionary changes. . . can be adequately accounted for in terms of the
Darwinian model. . .ĺ[369]
 |
Dr. Michael J. Dentonĺs Evolution: A Theory in Crisis was an influential critique of evolution by an agnostic scientist that contributed to the origins of Intelligent Design theory. |
- Introducing
his book Evolution Under the Microscope: A Scientific Critique of the
Theory of Evolution (Leighton Academic Press,
2002), research scientist David Swift writes:
ĹBecause evolution so permeates our education, it is not surprising that
from an early age I totally accepted the whole story of evolution from simple
forms of life, and probably of the origin of life itself. But in studying
for a degree in natural sciences ľ in which the theory of evolution formed
a backdrop to almost every aspect of the curriculum ľ I was prompted to
question whether the biochemical structures and mechanisms which were
being discovered at the time could really have arisen in an opportunistic
evolutionary manner: and it seemed to me that they could not. Over the
years since then, mostly spent as a research scientist, this view has been
strengthened as we have learned more of how biological systems work; and I
have become well aware that similar issues have been raised by others,
including experts in their field. Eventually, the opportunity arose to
research the subject more fully, and this book is the outcome. . . A
central issue is that, whilst I accept the principle of natural selection,
it cannot account for the formation of biological macromolecules, though
this is the usual evolutionary explanation for them. . . evolution has become
so generally accepted as a factual biological theory that the principle of
evolution has been assimilated by many other disciplines ľ such as
sociology, philosophy and even theology. I trust the book will be of
interest, indeed of value, to non-biologists from disciplines such as
these, who need to hear from a scientist of the substantial difficulties
with the theory ľ that it is nothing like so secure as they have probably
been led to believe.ĺ[370]
In response to a
recent American television series on evolution, 132 qualified scientists signed
a joint statement saying: ĹWe are sceptical of claims for the ability of random
mutation and natural selection to account for the complexity of life.ĺ[371]:
Signers of the statement questioning Darwinism came from throughout
the US and from several other countries, representing biology, physics,
chemistry, mathematics, geology, anthropology and other scientific fields.á
Professors and researchers at such universities as Princeton, MIT, U Penn, and
Yale, as well as smaller colleges and the National Laboratories at Livermore, CA and Los Alamos, N.M., are included.[372]
As of October
2003 over 300 scientists had signed the sceptical statement.[373]
Physicist
Paul Davies moved from promoting atheism in 1983 to realizing in 1984 that Ĺthe
laws [of physics] seem themselves to be the product of exceedingly ingenious
designĺ[374],
and affirming by 1989 that this fact is Ĺpowerful evidence that there is
something going on behind it all. The impression of design is overwhelming.ĺ[375] In his recent book, The
Origin of Life, Davies writes: ĹWhen I set out to write this book I was
convinced that science was close to wrapping up the mysteries of lifeĺs origin.
. . Having spent a year or two researching the field I am now of the opinion
that there remains a huge gulf in our understanding. . . This gulf in
understanding is not merely ignorance about certain technical details, it is a
major conceptual lacuna.ĺ[376]
While Davies says he is Ĺnot suggesting that lifeĺs origin was a supernatural
eventĺ[377],
he does think Ĺthat we are missing something very fundamental about the whole
businessĺ[378]
and confesses: ĹMy personal belief, for what it is worth, is that a fully
satisfactory theory of the origin of life demands some radically new ideas.ĺ[379], because Ĺscientists are
currently stumpedĺ.[380]
 |
In The Origin of Life (Penguin, 2003) Professor Paul Davies argues himself out of the view that science is close to wrapping up the mystery of lifeĺs origin and suggests that Ĺwe are missing something very fundamental about the whole businessĺ that will require some Ĺradically new ideasĺ. |
Time to Shift Paradigm?
ĹDarwinism is
too small to fit the facts it claims to explain, and ID is large enough to
include a modified form of Darwinism.ĺ ľ Benjamin Wiker[381]
Philosopher of
science Thomas Kuhn famously argued that major advances in science have
occurred through a succession of Ĺrevolutionsĺ wherein data accumulates that
does not fit with the established paradigm of Ĺnormal scienceĺ. At first there
are, quite naturally, vigorous attempts to absorb this new information into the
accepted paradigm: Ĺbut eventually the day comes when the paradigm can no
longer stand the burden of its inner contradictions, and a new paradigm is
established that makes sense of the new data in a more satisfactory way.á This
ôrevolutionö in paradigm shifts then makes way for a new period of ônormal
scienceö.ĺ[382]
Design theorists believe that there is a need to subsume Darwinĺs theory of
evolution within the context of a scientific paradigm of intelligent design,
because ĹScientific evidence, when evaluated without an overwhelming bias
toward materialism, does not support the Darwinian creation story that has
effectively become a state-sponsored religion in modernist culture. On the
contrary, the evidence actually supports the supposedly discredited view that
an intelligent designer. . . had to be involved in biological creation.ĺ[383]
Thomas Woodward argues that Darwinian sceptics like Michael Denton
and Phillip Johnson were pre-revolutionary in that they Ĺattacked Darwinism by
piling up scientific anomalies and analysing the underlying philosophical
assumptionsĺ, whereas ĹBehe and Dembski have made design theory revolutionary
in a Kuhnian sense by proposing a new heuristic paradigm that organizes
research in a different direction.ĺ[384]
Darwin found himself trapped between seeing everything in the
biological world as the result of direct and immediate intelligent design, or
seeing everything as the result of an unintended evolutionary process:
ĹI am conscious that I am in an utterly hopeless muddle. I cannot think that
the world, as we see it, is the result of chance; and yet I cannot look at each
separate thing as the result of Design.ĺ[385]
One way out of Darwinĺs muddle is to hypothesise that life is the result of a
combination of direct, immediate design and of an indirect, mediated
process of intended evolution. This conclusion, I believe, is the best
explanation of the available evidence. Darwin considered just such a hypothesis
in The Origin of Species, but rejected it because, as he said, its
proponents Ĺdo not pretend that they can define, or even conjecture, which are
the [directly] created forms of life, and which are those produced by secondary
laws. They admit variation as a vera causa in one case, they arbitrarily
reject it in another, without assigning any distinction in the two cases.ĺ[386] ID provides a way out of Darwinĺs muddle while satisfying the implicit demand of Darwinĺs objection to limiting the
explanatory work done by secondary causes.á Dembskiĺs work on CSI allows design
theorists to distinguish nonarbitrarily between what secondary causes
can and canĺt accomplish. Beheĺs concept of Ĺirreducible complexityĺ performs
the same service, albeit in a more limited fashion. As Dembski writes:
Prior to the rise of modern science all the emphasis was on
teleological guidance (typically in the form of divine design). Now the
pendulum has swung to the opposite extreme, and all the emphasis is on natureĺs
autonomy. . . Where is the point of balance that properly respects both, and in
which design becomes empirically evident? The search for that balance-point
underlies all design-theoretic research. Itĺs not all design or all nature but a
synergy of the two. Unpacking that synergy is the intelligent design research
program in a nutshell.[387]
ID doesnĺt hold that the neo-Darwinist theory of evolution is wholly
and straightforwardly wrong; but it does suggest that it is only a
partially successful theory that needs to be subsumed by a wider explanatory
hypothesis of design, somewhat as Newtonĺs Laws were subsumed by
Einsteinĺs: ĹIn the heady early days of Newtonian mechanics,ĺ notes Dembski,
Ĺphysicists thought Newtonĺs laws provided a total account of the constitution
and dynamics of the universe.á Maxwell, Einstein and Heisenberg each showed
that the proper domain of Newtonian mechanics was far more constricted. So,
too, the proper domain of the mutation-selection mechanism is far more constricted
than most Darwinists would like to admit.ĺ[388]
As Benjamin Wiker suggests: Ĺthe application of materialist principles to
nature has been extraordinarily fruitful, insofar as it represents a
half-truth, yet it has ended ultimately in undermining its own presuppositions.ĺ[389] Putting an interesting spin
on the matter, Joshua A. Smart writes: ĹID is in some sense an evolutionary
theory. It is a theory about when evolution is inadequate.ĺ[390]
Dembski acknowledges that Ĺthe Darwinian selection mechanism
constitutes a fruitful idea for biologyĺ[391]
He also accepts: Ĺthat some speciation occurs in the manner described by Darwin. . .ĺ[392] However: ĹDarwinism is the
totalising claim that this mechanism accounts for all the diversity of life.
The evidence simply does not support this claim. What evidence there is
supports limited variation within fixed boundaries, or what is typically called
microevolution.ĺ[393]
Moreover: ĹMacroevolution ľ the unlimited plasticity of organisms to diversify
across all boundaries ľ even if true, cannot legitimately be attributed to the
mutation-selection mechanism. To do so is to extrapolate the theory beyond its
evidential base.ĺ[394]
Dembski affirms the existence of a Ĺconsiderable overlapĺ[395] between Neo-Darwinism and
Intelligent Design, and points out that: Ĺintelligent design. . . does not
require that every aspect of biology be designed and is fully capable of
assimilating the Darwinian mechanism.ĺ[396]
As Wiker observes: ĹID proponents. . . do not deny many of the marvellous
things that Darwinism has uncovered, and so an ID account of biology would
include much of what Darwinists have discovered. What they question, however,
is the Darwinian assertion that such things are explicable solely as the result
of. . . unguided mechanisms.ĺ[397]
A Fruitful Scientific Research Program
Ĺthe ID approach
is both quite natural and scientifically fruitful.ĺ ľ Benjamin Wiker[398]
Dawkins avers
that: ĹWithout evolution, biology is a collection of miscellaneous facts.ĺ[399] Even if he were right about
this, wouldnĺt having a miscellaneous collection of facts be better than
having the facts distorted by a false theoretical framework?á Similarly,
Theodosius Dobzhansky propounded the maxim: ĹNothing in biology makes sense
except in the light of evolution.ĺ[400]
But as Jonathan Wellĺs responds, ĹA true scientist would say that nothing in
biology makes sense except in the light of evidence.ĺ[401]
Dawkinsĺ fear is doubly unfounded, because ID represents an
alternative theoretical framework for biology, a fruitful new Ĺparadigmĺ or
Ĺresearch programmeĺ. The ID approach is already paying dividends: ĹDesign is
not a science stopper. Indeed, design can foster inquiry where traditional
evolutionary approaches obstruct it.ĺ[402]
For example, Wiker points out that: ĹSince the last half of the 20th
century, the discovery of fine-tuning has been the impetus leading to the
discovery of more fine-tuningĺ.[403]
Wiker concludes: ĹBecause the universe is irreducibly complex ľ not just at one
level, but on successive layers of interrelated complexity ľ accepting the
apparent complexity as real does not eliminate science but destroys materialist
reductionism so that science may be released from the desire to shrink the
universe and instead may inquire ever more deeply into creation. . .ĺ[404] Scott Minnich, Associate
Professor of Microbiology at the University of Idaho, is an expert on the
flagellum (which he has studied for over fifteen years) who says that belief in
design has given him many research insights.[405]
Dembski concludes: ĹReinstating design within science can only enrich science.
. . By eschewing design, science has for too long operated with an inadequate
set of conceptual categories.á This has led to a constricted vision of reality,
skewing how science understands not just the world but also ourselves.ĺ[406] Woodward likewise welcomes ID
as a liberating paradigm:
To be jarred awake to the powerful role that metaphysical
commitments play in cosmology ľ especially their buttressing and protecting the
Darwinian knowledge-claims ľ can be upsetting and unpleasant, but the cognitive
health benefits are enormous. This awakening to the power of hidden
metaphysical foundations is the most inevitable and immediately beneficial sort
of ôarousing from dogmatic slumberö that Design could achieve. Through such a
rude awakening, the power of reason is newly energized. Indeed, such basic
notions as rationality, intelligence, and academic freedom are given new
impetus, a new breath of life. . .[407]
Darwinists once argued Ĺthat non-protein coding DNA are relics of
once-functioning genes or useless ôjunkö DNA that strongly argued against
design of the genome. . .ĺ[408]
However, Ĺnew research is beginning to overturn the view that most of the
genome has no functionĺ[409],
and this is a result that verifies ID in that, unlike Darwinism, ID naturally
suggests the hypothesis that so called Ĺjunk DNAĺ will turn out to have a
useful purpose:
huge stretches of genetic material dismissed in biology classrooms
for generations as Ĺjunk DNAĺ actually contain instructions essential for the
growth and survival of people and other organisms. . . scientists said the new
discoveries were likely to force them to abandon the term Ĺjunk DNAĺ and send
them back to the drawing board to come up with sweeping new models for how
nature builds and maintains organisms.[410]
Dembski argues:
ĹID theorists should be at the forefront in unpacking the information contained
within biological systems. If these systems are designed, we can expect the
information to be densely packed and multi-layered (save where natural forces
have attenuated the information). Dense, multi-layered embedding of information
is a prediction of ID.ĺ[411]
A similar shift of opinion verifying the design hypothesis has occurred in the
case of so called vestigial structures: Ĺmost of the structures regarded as
vestigial in humans a hundred years ago are now known to have a function. . .ĺ[412] Here again: Ĺwe find design
encouraging scientists to look for function where evolution discourages it.ĺ[413]
A Fledgling Breakthrough
As Jonathan
Wells says, ID is Ĺa fledgling program, but there are some promising leads. It
could have saved us 25 years if an ID approach to ôjunkö DNA had been pursued
back when it was discovered. There are also applications now being pursued
actively in embryology and the study of bacteria.ĺ[414] Further conceptual and
experimental work certainly remains to be done (after all, thatĺs the whole
point of a scientific research programme)[415];
and only time will tell if ID can succeed in prompting a paradigm shift within
the natural sciences. Whether or not it does so depends upon overcoming
negative philosophical assumptions as much as upon matters of empirical
evidence (indeed, it depends upon getting people to approach and evaluate the
empirical evidence without a philosophical bias against design). As Dembski
writes: ĹIĺll be the first to admit that intelligent design is an ambitious
program and that it may not pan out. But it needs first to be fairly
discussed.ĺ[416]
Paul Davies comments: ĹDembskiĺs attempt to quantify design, or provide
mathematical criteria for design, is extremely useful.á Iĺm concerned that the
suspicion of a hidden agenda is going to prevent that sort of work from
receiving the recognition it deserves. Strictly speaking, you see, science
should be judged purely on the science and not on the scientist.ĺ[417] According to mathematician
Wolfgang Smith, a Professor Emeritus at Oregon State University:
The intelligent design theory is the first rigorous refutation of
Darwinism on a scientific and mathematical basis. This impresses me as a major,
scientific breakthrough which, hopefully in time, will be recognized by the
scientific community at large.[418]
Indeed, despite
the deep-rooted naturalistic prejudices of the reigning paradigm of Ĺnormal
scienceĺ, ID research Ĺis being published and cited in the peer-reviewed
scientific literature (including the biological literature)ĺ[419], appearing in such journals
as: Proceedings of the
National Academy of Sciences, Journal of
Molecular Biology, Protein Science, Journal of Theoretical
Biology, Origins of Life and Evolution of the Biosphere, Proceedings of the Biological Society of Washington and Annual Review of Genetics.
While many scientists and philosophers (including Christian
scientists and philosophers) continue to believe in evolution, many scientists
and philosophers (including Christian scientists and philosophers) believe that
the neo-Darwinian paradigm is seriously flawed. Huston Smithĺs arresting
verdict may prove to be prophetic: ĹDarwinism is in fact dying, and its death
signals the close of our age.ĺ[420]
Recommended Resources
Access Research
Network @ www.arn.org
Axe, Douglas, ĹExtreme Functional Sensitivity to Conservative Amino Acid Changes on Enzyme
Exteriorsĺ @ www.mrc-cpe.cam.ac.uk/publications/axe_jmb_2000.pdf
Behe,
Michael J., ĹMolecular Machines: Experimental Support for the Design Inferenceĺ @ www.arn.org/docs/behe/mb_mm92496.htm
Behe, Michael J., ĹBlind Evolution or Intelligent Design?ĺ @ www.discovery.org/scripts/viewDB/index.php?command=view&program=CSC%20-%20Scientific%20Research%20and%20Scholarship%20-%20Science&id=1205
Berlinski,
David, ĹThe Deniable Darwinĺ @ www.discovery.org/scripts/viewDB/index.php?command=view&id=130
Craig, William Lane, ĹReview: The Design Inference ľ Eliminating chance through small
possibilitiesĺ @ www.leaderu.com/offices/billcraig/docs/design.html
William
A. Dembskiĺs Design Inference Website @ www.designinference.com/
Dembski,
William A., ĹIntelligent Design as a Theory of Informationĺ @ www.arn.org/docs/dembski/wd_idtheory.htm
Dembski,
William A., ĹThe Third Mode of Explanationĺ @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=61
Dembski,
William A, ĹThe Logical Underpinnings of Designĺ @ www.designinference.com/documents/2002.10.logicalunderpinningsofID.pdf
Dembski, William A., ĹIrreducible Complexity
Revisitedĺ @ www.designinference.com/documents/2004.01.Irred_Compl_Revisited.pdf
Meyer,
Stephen C., ĹDNA and Other Designsĺ @ www.arn.org/docs/meyer/sm_dnaotherdesigns.htm
Meyer,
Stephen C., ĹThe Origin of Biological Information and the Higher Taxonomic
Categoriesĺ, @ www.discovery.org/scripts/viewDB/index.php?command=view&id=2177&program=CSC%20-%20Scientific%20Research%20and%20Scholarship%20-%20Science
Meyer,
Stephen C., et al, ĹThe Cambrian Explosion: Biologyĺs Big Bangĺ @ www.discovery.org/articleFiles/PDFs/Cambrian.pdf
Minnich, Scott A. & Stephen C. Meyer, ĹGenetic Analysis of Coordinate Flagellar and Type II
Regulatory Circuits in Pathogenic Bacteria ĺ @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=148
Moreland, J. P., ĹIs Science a Threat or a Help to Faith? A Look at
the Concept of Theistic Scienceĺ @ www.afterall.net/index.php/papers/18
Moreland, J. P., ĹComplimentarity, Agency Theory, and the
God-of-the-Gapsĺ @ www.afterall.net/index.php/papers/490579
Moreland J. P., ĹScientific Creationism, Science, and Conceptual
Problemsĺ @ www.afterall.net/index.php/papers/490578
Newman, Robert C. et al, ĹThe Status of Evolution as a Scientific Theoryĺ @ www.arn.org/docs/newman/rn_statusofevolution.htm
Plantinga, Alvin, ĹMethodological Naturalism?ĺ @ http://id-www.ucsb.edu/fscf/library/plantinga/mn/home.html
Plantinga, Alvin, ĹWhen Faith and Reason Clash: Evolution and the
Bibleĺ @ www.asa3.org/ASA/dialogues/Faith-reason/CRS9-91Plantinga1.html
Plantinga, Alvin, ĹEvolution, Neutrality, and Anticedent
Probability: a Reply to Van Till and McMullenĺ @ www.asa3.org/ASA/dialogues/Faith-reason/CRS9-91Plantinga2.html
Plantinga, Alvin, ĹOn Rejecting the Theory of Common Ancestry: A
Reply to Haskerĺ @ www.asa3.org/ASA/dialogues/Faith-reason/PSCF12-92Plantinga.html
Wells, Jonathan, ĹSurvival
of the Fakestĺ @á www.discovery.org/articleFiles/PDFs/survivalOfTheFakest.pdf
Williams, Peter S., ĹA Rough Guide to Creation and Evolutionĺ @ www.arn.org/docs/williams/pw_roughguidetocreationandevolution.htm
Williams,
Peter S., ĹDarwinsĺ Rottweiler & the Public Understanding of Scientismĺ @ www.arn.org/docs/williams/pw_dawkinsfallacies.htm
Williams,
Peter S., ĹReviewing the Reviewers: Pigliucci et al on ôDarwinĺs Rottweiler & the public understanding of scienceöĺ @ www.arn.org/docs/williams/pw_pigliucci_reviewingreviewers.htm
Williams,
Peter S., ĹDesign Arguments: Amazon Listmania Listĺ @ www.amazon.com/exec/obidos/tg/listmania/list-browse/-/3CKOYI2H94VQU/ref=cm_aya_av.lm_more/002-9805224-1419262
Beckwith,
Francis J., Law, Darwinism, and Public Education: The Establishment
Clause and the Challenge of Intelligent Design, (New York: Rowman & Littlefield, 2003)
Behe, Michael J., Darwinĺs Black Box, (Free Press, 1998)
Behe,
Michael J., et al, Science and Evidence for Design in the Universe,
(San Francisco: Ignatius, 2000)
Campbell, John Angus & Stephen C. Meyer (ed.ĺs), Darwinism, Design, And Public
Education, (East Lansing: Michigan State University Press, 2003)
Dembski,
William A., The Design Revolution: Answering the Toughest Questions about
Intelligent Design, (IVP, 2004)
Dembski, William A., No
Free Lunch: Why Specified Complexity Cannot be Purchased without Intelligence,
(Lanham: Rowman & Littlefield, 2001)
Dembski,
William A. (ed.), Uncommon Dissent: Intellectuals Who Find Darwinism
Unconvincing, (Wilmington, Del: ISI Books, 2004)
Dembski,
William A. and James M. Kushner (ed.ĺs), Signs of Intelligence, (Grand Rapids , Michigan: Brazos Press, 2001)
Johnson,
Philip E., Darwin on Trial, (Downers Grove: IVP)
Moreland,
J.Pá (ed.), The Creation Hypothesis, (Downers Grove, Illinois: IVP,
1994)
Overman,
Dean L., A Case Against Accident and Self-Organization, (Rowman & Littlefield, 1997)
Ratzsch, Del, Science & Its Limits: The Natural Sciences in Christian Perspective
(Leicester: Apollos, 2000)
Strobel,
Lee, The Case for a Creator, (Zondervan, 2004)
Thaxton,
Charles B. et al, Mystery of Lifeĺs Origins: Reassessing Current
Theories, (Ashgate, 1987)
Wells,
Jonathan, Icons of Evolution, (Washington: Regnery Publishing, 2000)
Williams,
Peter S., I Wish I Could Believe in Meaning: A Response to Nihilism,
(Damaris, 2004)
Woodward,
Thomas, Doubts About Darwin: A History of Intelligent Design, (Grand Rapids: Baker, 2003)
On
Video or DVD: Unlocking
the Mysteries of Life (Illustrated Media)
Endnotes
[1] Beckwith, Francis J, Law, Darwinism, and Public
Education: The Establishment Clause and the Challenge of Intelligent Design,
(New York: Rowman & Littlefield, 2003), p. xiii.
[2] cf. Craig, William Lane, ĹReview: The Design
Inference ľ Eliminating chance through small possibilitiesĺ @ www.leaderu.com/offices/billcraig/docs/design.html;
Dembski, William A., ĹThe Third Mode of Explanationĺ @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=61;
Dembski, William A., ĹIntelligent Design as a Theory of Informationĺ @ www.arn.org/docs/dembski/wd_idtheory.htm;
Dembski, William A, ĹThe Logical Underpinnings of Designĺ @ www.designinference.com/documents/2002.10.logicalunderpinningsofID.pdf;
Dembski, William A., No Free Lunch: Why Specified Complexity Cannot be
Purchased without Intelligence, (Lanham: Rowman & Littlefield, 2001)
[8] cf. Stephen C. Meyer, ĹThe Origin of Biological
Information and the Higher Taxonomic Categoriesĺ, @ www.discovery.org/scripts/viewDB/index.php?command=view&id=2177&program=CSC%20-%20Scientific%20Research%20and%20Scholarship%20-%20Science;
Kurt P. Wise, ĹThe Origin of Lifeĺs Major Groupsĺ, in J.P. Moreland (ed.), The
Creation Hypothesis, (IVP, 1994); Robert F. Dehaan and John L. Wiester, ĹThe Cambrian Explosion: The Fossil Record and Intelligent Designĺ, in William
A. Dembski & James M. Kushiner (ed.ĺs), Signs of Intelligence,
(Brazos Press, 2001); & Stephen C. Meyer, Marcus Ross, Paul Nelson and Paul
Chien, ĹThe Cambrian Explosion: Biologyĺs Big Bangĺ, in John Angus Campbell
& Stephen C. Meyer (ed.ĺs), Darwinism, Design, And Public Education,
(Michigan State University Press, 2003) @ www.discovery.org/articleFiles/PDFs/Cambrian.pdf
[9] Beckwith, op
cit, p. xvi-xvii.
[12] Dawkins, A
Devilĺs Chaplain, p. 219.
[13] Krishtalka,
Leonard, quoted by Glenn Branch in ĹHuman Nature After Darwin by Janet Radcliffe
Richardsĺ, Philosophy Now, 40, March/April 2003, p. 44.
[15] Gould, Stephen
Jay, Natural History, March 1997, quoted Woodward, op cit, p.
210.
[16] ARN guide to Evolution,
op cit.
[17] John Angus Campbell, ĹWhy Are We Still Debating Darwinism?,
Campbell & Meyer (ed.ĺs), Darwinism, Design, And Public Education,
(Michigan University Press, 2003), p. xxvi.
[18] ĹIntelligent
design: The New ôBig Tentö for Evolutionĺs criticsĺ, University of Wisconsin ľ Madison News, February 19, 2004.
[19] Wiker, Moral
Darwinism, p. 77.
[20] Woodward, op
cit, p. 196, my italics.
[21] Dembski, ĹWhat
Every Theologian Should Know about Creation, Evolution, and Designĺ, op cit.
[22] Johnson,
Phillip E., Darwin on Trail, (Downers Grove, IVP, 19932), p. 14á & 115.
[23] Woodward,
Appendix 2, op cit, p. 223.
[24] Johnson, Reason
in the Balance, (Downers Grove: IVP, 1995), p. 74.
[26] Beckwith, op
cit, p. 153.
[27] Woodward,
Thomas, Doubts about Darwin: A History of Intelligent Design, (Grand Rapids: Baker, 2003), p. 10 & 20.
[29] Beckwith, op
cit, p. 164.á
[31] Behe, Michael
J., ĹThe Modern Intelligent Design Hypothesisĺ, Philosophia Christi,
Series 2, Volume 3, Number 1, 2001, p. 165.
[34] Dembski, ĹIs
Intelligent Design a Form of Natural Theology?ĺ, op cit.
[35] ARN guide to Evolution.
[36] Wiker, op
cit, p. 77.
[37] Nichols,
Terence, The Sacred Cosmos, (Grand Rapids: Brazos Press, 2003), p. 61.
[38] Plato, The Laws, book X.
[40] cf. Plantinga, Alvin, ĹMethodological
Naturalism?ĺ @ http://id-www.ucsb.edu/fscf/library/plantinga/mn/home.html;
Craig, William Lane and Moreland, J. P., Philosophical Foundations for a
Christian Worldview (Downers Grove: IVP, 2003); Moreland, J.Pá (ed.), The
Creation Hypothesis, (Downers Grove, Illinois: IVP, 1994)
[41] Dawkins, Richard, The Blind Watchmaker, p. 151, my emphasis.
[42] Lewontin, Richard, ĹBillions and Billions of Demonsĺ, New York Review of Books, January 9, 1997.
[43] Dembski, William A., ĹWhat Every Theologian Should Know about
Creation, Evolution and Designĺ, p. 6, my italics.
[44] Ratzsch, Del, Science & Its Limits, (Leicester: Apollos,
2000), p. 123-124.
[45] Witham, Larry, By Design, (San Francisco: Encounter Books,
2003), p. 143.
[47] DeWolf et al, ĹTeaching the Controversy: Is it Science,
Religion, or Speech?ĺ, John Angus Campbell & Stephen C. Meyerĺs (ed.ĺs), Darwinism,
Design, And Public Education, (Michigan State University Press, 2003), p. 79.
[48] Moreland, J.P., ĹTheistic Science & Methodological Naturalismĺ,
Moreland (ed.), The Creation Hypothesis, p. 55.
[50] Beckwith, op
cit, p. 107.
[51] Michael Ruse, Can a Darwinian be a Christian?, p. 101.
[52] Michael Ruse, quoted by John Angus Campbell, ĹIntelligent Design, Darwinism,
and Public Education Philosophyĺ, in Campbell & Meyer (ed.ĺs), Darwinism,
Design, And Public Education, (Michigan State University Press, 2003), p.
30.
[53] John Angus Campbell, op cit, p. xii.
[54] Dembski, Mere
Creation, (Downers Grove: IVP, 1996), p. 17.
[55] Dembski, ĹThree
Frequently Asked Questions About Intelligent Designĺ, op cit.
[56] Dembski, op
cit, p. 17.
[61] Bocarsly, Andrew and Kaita, Robert, in Woodward, op. cit.,
p.212.
[62] Meyer, Steven C., ĹThe Explanatory Power of Designĺ in William A.
Dembski (ed.), Mere Creation (Downers Grove: IVP, 1998).
[63] Dawkins, Richard, Climbing Mount Improbable (Viking, 1996),
p.259.
[64] Dawkins, Richard, ĹDarwinĺs Dangerous Discipleĺ at
www.skeptic.com/03.4.miele-dawkins-iv.html
[65] Dawkins, Richard, A Devilĺs Chaplain (London: Weidenfeld & Nicholson, 2003), p.248.
[66] Dawkins, ĹDarwinĺs Dangerous Discipleĺ at
www.skeptic.com/03.4.miele-dawkins-iv.html
[67] Wiker, Benjamin, op. cit.
[68] Wald, George,ĹInnovation and Biologyĺ, Scientific American,
199 (Sept. 1958): 100.
[69] Johnson, Phillip E., ĹPositional Paper on Darwinismĺ, in Woodward,
Appendix 2, op. cit., p.219.
[70] Kerkut, G. A., Implications of Evolution (Pergamon Press),
p.150, quoted by John Blanchard, in Does God Believe in Atheists? (Darlington: Evangelical Press, 2000), p. 297.
[72] Johnson, Phillip E., ĹWhat is Darwinismĺ, Objection Sustained,
(Downers Grove: IVP, 1998), p. 33.
[73] Craig, William Lane, Philosophy of Religion: A Reader and Guide
(Edinburgh University Press, 2002), p.72.
[77] The simulated monkeys typed: Ĺ1. Citizen. Before
wZgJ
8GPxwFnwvG&iX4tKfo("2ny!3Pp. . .ĺ which matched: Ĺ1. Citizen. Before
we proceed any further, heare me speake. . .ĺ Interestingly, it took 2,182,280,000 billion billion
monkey-years to produce 18 letters from ĹTimon of Athensĺ (ĹPoet. Good day
SirBf4I.ö ôM 'O.)jdtQ4Q].IPRe&BSJF. . .ĺ matched ĹPoet. Good day
Sir Pain. I am glad y'are well . .) & 42,162,500,000 billion
billion monkey-years to produce 19 letters from ĹTwo Gentlemen of Veronaĺ
(ĹVALENTINE. Cease to1d0r:eFLP0FRjWK78aXzVOwm)-';8.t. . .ĺ matched: ĹVALENTINE.
Cease to persuade, my loving Proteus. . .ĺ)
[78] Dembski, Intelligent
Design, (Downers Grove: IVP, 1999), p. 166.
[79] Dawkins, The
Blind Watchmaker, (London: Penguin, 1988), p. 139.
[80] Dembski, No
Free Lunch, (New York: Rowman & Littlefield, 2002), p. 157.
[83] Dawkins, Climbing Mount Improbable, (TSP: Viking, 1996), p.
4
[86] Dawkins, The Blind Watchmaker, op cit, Preface, p. x.
[87] ibid, p. 4-5.á This statement embodies the false dilemma
that either design or evolution is true, when these options are in fact
logically compatible. cf. Stephenĺs Guide to the Logical Fallacies, ĹFalse Dilemmaĺ @ www.intrepidsoftware.com/fallacy/fd.php
(05/04/2004)
[92] Keith, Alan, Philosophy Now, Issue 45, March/April 2004,
Letters, p. 42.
[93] cf. Williams,
Peter S., ĹDarwinĺs Rottweiler & the public understanding of scienceĺ, Philosophy
Now Issue 44 (Jan/Feb 2004)
[94] Massimo
Pigliucci is associate Professor at the University of Tennessee in Knoxville, where he teaches ecology and evolutionary biology.á Professor Pigliucci has a
PhD in botany from the University of Connecticut and a PhD in philosophy from
the University of Tennessee. Pigliucci has published many technical papers and
two books on evolutionary biology (Phenotypic Evolution, with Carl
Schlichting, and Phenotypic Plasticity: Beyond Nature vs. Nurture.)á He
has been awarded the Oak Ridge National Labs award for excellence in research
(several times), and has won the prestigious Dobzhensky prize from the Society
for the Study of Evolution (of which he is now vice president). A self-styled
Ĺskepticĺ, Pigliucciĺs articles have appeared in: The Skeptic, Free
Inquiry, Philosophy Now and The Philosophers Magazine. cf. www.infidels.org/library/modern/massimo_pigliucci/index.shtml
[95] Pigliucci,
Massimo, et al, ĹThe Alleged Fallacies of Evolutionary Theoryĺ,
published in Philosophy Now Issue 46 (May/June 2004).
[100] Koons, Robert
C., ĹThe Check Is In The Mail: Why Darwinism Fails to Inspire Confidenceĺ, in
Dembski, William A., (ed.), Uncommon Dissent, (2004), p. 7.
[106] Dawkins, op cit, p. 3.
[113] Dawkins, op cit, p. 3.
[114] Richard Dawkins, in interview with Keith Ward, The Turn of The
Tide, (BBC, 1986), p. 28, my italics.
[115] Behe, ĹBlind
Evolution or Intelligent Design?ĺ
[116] Stephen Hawking, quoted by John Boslough, Masters of Time ľ Cosmology at the End of Innocence, (New York: Addison-Wesley Publishing
Company, 1992), p. 55.
[118] Dawkins, The Blind Watchmaker, op cit.
[119] Adams, Douglas, The Salmon of Doubt (Pan Books, 2003), p.131.
[120] Overman, A
Case Against Accident and Self-Organization (Rowman & Littlefield,
1997), p. 128.
[121] Moreland, J.
P.á and Craig, William Lane, Philosophical Foundations For A Christian
Worldview, p.487.
[124] J.P. Moreland, Scaling the Secular City, (Grand Rapids:
Baker, 1987).
[125] cf. Stephen C. Meyer, ĹEvidence for Design in Physics and Biologyĺ
in Michael Behe, William A. Dembski & Stephen C. Meyer, Science and
Evidence for Design in the Universe, (San Francisco, Ignatius, 2000), p. 60.
[126] cf. Jimmy H. Davies and Harry L. Poe, Designer Universe, (Nashville: Broadman & Holman, 2002), p. 85.
[127] Penrose, Roger, The Emperorĺs New Mind, (New York, Oxford,
1989), p. 344.
[128] Pageĺs calculation is quoted by L. Stafford Betty and Bruce
Cordell, ĹThe Anthropic Teleological Argumentĺ, International Philosophical
Quarterly 27, no. 4., (December 1987).
[129] Fred Hoyle, as quoted in Fred Hereen, Show Me God, (Search
Light Publishing, 1995), p. 179.
[130] Clark, Stephen R. L., God, Religion and Reality (London: SPCK, 1998), p.106.
[131] Dembski, op
cit, p. 16-17.
[132] Moreland, J.P. & William Lane Craig, Philosophical Foundations For A Christian
Worldview, (Downerĺs Grove: IVP, 2003), p. 62.
[133] Dawkins, The Blind Watchmaker, op cit, p. 9.
[134] Dawkins, ĹDarwinĺs Dangerous Discipleĺ at
www.skeptic.com/03.4.miele-dawkins-iv.html
[136] Philosophy
Now, Issue 47, August/September 2004, p. 22
[138] Asimov, Isaac, Please Explain, quoted by Alan Hayward, in Does
God Exist? Science says ĹYesĺ (Basingstoke: Lakeland, 1979), p.76.
[139] Hayward, op cit., p.76.
[140] Wiker, Benjamin, ĹDoes Science Point to God?ĺ at www.arn.org/docs2/news/doessciencepointtogod040903.htm
[141] Woodward, Thomas, Doubts About Darwin (Grand Rapids: Baker,
2003), p.44.
[142] Overman, Dean L., A Case Against Accident and Self-Organization (Rowman & Littlefield, 2001), p.41.
[143] Overman, op. cit., p. 41.
[144] Denton, Michael, Evolution: A Theory in Crisis (Woodbine
House, 1996), p.262.
[145] Varghese, The
Wonder of the World, op cit, p. 276.
[146] Yockey, Hurbert P., ĹA calculation of the Probability of
Spontaneous Biogenesis by Information Theory,ĺ Journal of Theoretical
Biology, 67, (1977), p. 377.
[147] Overman, op. cit., p. 40.
[148] Denyse
OĺLeary, By Design or by Chance? The Growing Controversy on the
Origins of Life in the Universe, (Minneapolis: Ausberg Books, 2004), p. 8.
[149] Beadle, George, ĹThe Language of the Geneĺ in The Language of
Science (New York: Basic Books, 1963), p.62.
[150] Delbruck, Max, interview in Horace Freeland Judson, The Eighth
Day of Creation (New York: Simon and Schuster, 1979), p.213.
[151] Blanchard, John, Does God Believe in Atheists? (Darlington: Evangelical Press, 2000), p. 292.
[152] Varghese, op
cit, p. 276.
[153] Quoted by
Varghese, The Wonder of the World, p. 278.
[154] Sarfati, Jonathan, Refuting Evolution 2 (Master Books,
2003), p.58.
[155] Bradley, Walter L., in Strobel, The Case for Faith, op.
cit., p.111. Bradley is using Ĺfaithĺ in the same sense that Dawkins uses
the term to mean belief without or against evidence.
[156] Dawkins, Richard, Climbing Mount Improbable (Viking, 1996),
p.259.
[157] Dawkins, Richard, ĹDarwinĺs Dangerous Discipleĺ at
www.skeptic.com/03.4.miele-dawkins-iv.html
[158] Dawkins, Richard, A Devilĺs Chaplain (London: Weidenfeld & Nicholson, 2003), p.248.
[159] Dawkins is not alone in his leap of faith. For example, Harvard
biologist George Wald dismissed the design explanation for life but candidly
admitted: ĹI will not believe that philosophically because I do not want to
believe in God. Therefore, I choose to believe in that which I know is
scientifically impossible: spontaneous generation [that life arose from
non-living matter] arising to evolution.ĺ ľ ĹInnovation and Biologyĺ, Scientific
American, 199 (Sept. 1958): 100.
[160] Johnson, Phillip E., ĹPositional Paper on Darwinismĺ, in Woodward,
Appendix 2, op. cit., p.219.
[162] Bradley, Walter L., in Strobel, The Case for Faith
(Zondervan, 2000), p.100.
[163] Kerkut, G. A., Implications of Evolution (Pergamon Press),
p.150, quoted by John Blanchard, in Does God Believe in Atheists? (Darlington: Evangelical Press, 2000), p.297.
[164] Shapiro, Robert, Origins: A Skepticĺs Guide to the Creation of
Life on Earth (New York: Summit Books, 1986), p.112.
[165] Schidlowsky, Manfred, quoted by Overman, op. cit., p.43.
[167] Meyer, Steven C., ĹThe Explanatory Power of Designĺ in William A.
Dembski (ed.), Mere Creation (Downers Grove: IVP, 1998).
[168] Thaxton, Charles, Bradley, Walter and Olsen, Roger, The Mystery
of Lifeĺs Origin (Ashgate, 1987), p.185.
[169] Richard Dawkins, The Blind Watchmaker, (London: Penguin, 1988),
p. 139.
[170] Dawkins, op cit, p. 260.
[172] Ross, Hugh, ĹAstronomical Evidences for a Personal, Transcendent
Godĺ, in J. P. Moreland (ed.), The Creation Hypothesis (Downers Grove:
IVP, 1994), p.170.
[173] Faulkner, Danny R., in John F. Ashton (ed.), On the Seventh Day
(Green Forrest: Master Books, 2002), p.107.
[176] Ross, Hugh, ĹAstronomical Evidences for a Personal, Transcendent
Godĺ, op. cit., p.169-170.
[177] Ward, Peter and Brownlee, Don, Rare Earth (New York: Springer-Verlag,
2000), p.35, 37.
[178] Meyer, Stephen C., ĹEvidence for design in Physics and Biologyĺ, in Science and Evidence for Design in the Universe (San Francisco:
Ignatius, 2000), p.75.
[179] Dawkins, Richard, The Blind Watchmaker (London: Penguin,
1990), p.139.
[180] Wickramasinghe, Chandra, cited in Roy Abraham Varghese (ed.),
The Intellectuals Speak about God (Regnery Gateway), p.33, quoted by John
Blanchard, op. cit., p.298.
[181] Behe, Michael J., ĹThe Modern Intelligent Design Hypothesisĺ, Philosophia
Christi, Series 2, Volume 3, Number 1, 2001, p.177.
[183] Ward, Keith, God, Faith & The New Millennium (Oxford:
OneWorld, 1999), p.110.
[184] Hoyle, Fred and Wickramasinghe, Chandra, Evolution from Space
(Dent, 1981), p.24-148.
[186] Geisler, Norman L. and J. Kerby Anderson, Origin Science,
(Grand Rapids: Baker, 1987), p. 159 -164.
[188] Meyer, Stephen
C., ĹThe Origin of Biological Information and the Higher Taxonomic Categoriesĺ, Proceedings of the Biological Society of Washington (volume 117, no. 2,
2004, pp. 213-239).
[192] Zimmer, Carl, Evolution,
(New York: Harper Collins, 2001), p. 325.
[194] David Berlinksi, ĹBerlinksi replies to H. Allen Orrĺ, in Uncommon
Dissent, op cit, p. 286.
[195] Robert C. Newman et al, Whatĺs Darwin Got to Do With It?,
(IVP), p. 39.
[197] cf. Ahuja, Anjana, ĹA Species is Born: Scientists believe that they
may be witnessing evolution in action on our northern coastlineĺ, The Times,
T2, 22 July 2004, p. 14.á This report concerns two varieties or morphs of
Yorkshire periwinkles (mid-shore and high-shore) that may be in the
process of becoming separate species, in that they are currently not keen on
breeding with each other: ĹThe problem with watching just one freeze-frame [of
a presumed evolutionary history]. . . is not knowing what came before or what
will happen in the future.á For example, it is possible, though unlikely, that
the reverse is happening, and that two species are merging into one.ĺ
[198] David DeWolf, Stephen C. Meyer and Mark E. DeForrest, ĹTeaching the
Controversy: Is it Science, Religion, or Speech?ĺ, in John Angus Campbell &
Stephen C. Meyer (ed.ĺs), Darwinism, Design, And Public Education,
(Michigan State University Press, 2003), p. 66.
[199] Jonathan Wells, in Lee Strobel, The Case for a Creator,
(Zondervan, 2004), p. 43.
[200] Koons, op
cit, p. 4.
[201] Dehaan, Robert F. and John L. Wiester, ĹThe Cambrian Explosion: The
Fossil Record and Intelligent Designĺ, in William A. Dembski & James M.
Kushiner (ed.sĺ), Signs of Intelligence, (Brazos Press, 2001), p. 150.
[202] Kitts, David B., ĹPaleontology and Evolutionary Theoryĺ, Evolution,
vol. 28, 1974, p. 467.
[204] Sunderland, Luther, Darwinĺs Enigma, (Master Books, 1998),
p. 11.
[205] Ankerberg, John & John Weldon, ĹRational Inquiry & the
Force of Scientific Data: Are New Horizons Emerging?, in J.P. Moreland (ed), The
Creation Hypothesis, (IVP, 1994), p. 278.
[206] Niles Eldridge, The Myths of Human Evolution, (Columbia
University Press, 1982), p. 59.á The theory of Ĺpunctuated equilibriumĺ, with
which Eldridge is associated, Ĺis really more of an observation ľ based on the
fossil record, which shows appearance of new species (for example, in the
Cambrian explosion) than it is a theory in the usual senseĺ, and while Ĺthere
has been a major ongoing debate among evolutionists themselves on the likelihood
of the punctuated equilibrium model verses the traditional gradualistic oneĺ it
seems that Ĺthe traditional gradualism model still has strong supportĺ; and in
any case ĹWhichever model evolutionists choose to suggest. . . the lack of a
single example of a real transition is evidence enough that the fossil record
does not support evolution.ĺ (Ralph O. Muncaster, Dismantling Evolution,
p. 86-87.)á Phillip E. Johnson writes: ĹMost evolutionary biologists do not
accept Eldridge and Gouldĺs hypothesis that evolutionary change is closely
associated with speciation. . .á Speciation and change in form. . . seem to be
different phenomena. . .á For these and other reasons, orthodox neo-Darwinists
prefer to explain sudden appearance on the traditional basis of gaps in the
fossil record. . .ĺ (Darwin on Trial, p. 52-53.)á For a critique of both
Darwinian gradualism and punctuated equilibrium in relation to the fossil
record cf. Stephen C. Meyer, Marcus Ross, Paul Nelson and Paul Chien, ĹThe
Cambrian Explosion: Biologyĺs Big Bangĺ, @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=29 and in John Angus Campbell & Stephen C. Meyer (ed.ĺs), Darwinism,
Design, And Public Education, (Michigan State University Press, 2003), p.
338.
[207] David Raup, ĹConflicts between Darwin and Paleontologyĺ, Field Museum of Natural History Bulletin, January, 1979.
[208] Muncaster, op cit, p. 81.
[210] Johnson, Phillip E., ĹEvolution as Dogmaĺ, in Uncommon Dissent,
op cit, p. 26.
[211] Hunter, Cornelius G., ĹWhy Evolution Fails the Test of Scienceĺ, in Uncommon Dissent, op cit, p. 206.
[212] Eldridge, Niles, Harperĺs Magazine (February, 1985), p. 60.
[213] Stanley, Steve M., Macroevolution, p. 59, quoted by
Ankerberg & Weldon, op cit.
[214] Nelson, Gareth, quoted by Johnson, ĹEvolution as Dogmaĺ, op cit.
[215] Carroll, Robert, Patterns and Processes of Vertebrate Evolution,
(Cambridge University Press, 1997), p. 8.
[216] Hartwig, Mark, ĹChallenging Darwinĺs Mythsĺ, in James P. Gills
& Tom Woodward (ed.ĺs), Darwinism under the Microscope , p. 26.
[217] Quoted by Woodward, Tom, ĹOf Canadian Oddballs and Chinese
Monstersĺ, in James P. Gills & Tom Woodward (ed.ĺs), Darwinism under the
Microscope, (Charisma House, 2002), p. 105.
[218] Stephen C. Meyer, Marcus Ross, Paul Nelson and Paul Chien, ĹThe
Cambrian Explosion: Biologyĺs Big Bangĺ, @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=29 and in John Angus Campbell & Stephen C. Meyer (ed.ĺs), Darwinism,
Design, And Public Education, (Michigan State University Press, 2003), p.
338.
[219] Beckwith, Francis J., Law, Darwinism, And Public Education,
(Rowman & Littlefield, 2003), p. 118. cf. Kurt P. Wise, ĹThe Origin of
Lifeĺs Major Groupsĺ, in J.P. Moreland (ed.), The Creation Hypothesis,
(IVP, 1994); Robert F. Dehaan & John L. Wiester, ĹThe Cambrian Explosion:
The Fossil Record and Intelligent Designĺ, in William A. Dembski & James M.
Kushiner (ed.sĺ), Signs of Intelligence, (Brazos Press, 2001); &
Stephen C. Meyer, Marcus Ross, Paul Nelson & Paul Chien, ĹThe Cambrian
Explosion: Biologyĺs Big Bangĺ, in John Angus Campbell & Stephen C. Meyer
(ed.ĺs), Darwinism, Design, And Public Education, (Michigan State
University Press, 2003) & @ www.discovery.org/scripts/viewDB/filesDB-download.php?id=29
[221] McDonald,
J.F., ĹThe molecular basis of adaptation: a critical review of relevant ideas
and observationsĺ, Annual Review of Ecology and Systematics, 14:77-102,
1983, p. 93.
[226] Varghese, op
cit, p. 289.
[227] Johnson, Darwin on Trial, op cit, p. 27.
[229] Thompson,
Keith Stewart, ĹMacroevolution: The Morphological Problemĺ, American
Zoologist 32 (1992): 106-112.
[230] Hull, David L., Darwin and his Critics, p. 7, quoted by
Ankerberg & Weldon, op cit.
[231] Colson, Charles and Nancy Pearcey, Developing a Christian
Worldview of Science And Evolution, (Tyndale House, 2001), p. 73.
[232] Hunter, Cornelius G., Darwinĺs Proof, (Brazos Press, 2003),
p. 60.
[233] Dehaan and Wiester, op cit, p. 150.
[235] Hunter, ĹWhy Evolution Fails the Test of Scienceĺ, in Uncommon
Dissent, op cit, p. 203.
[236] Wells, in Strobel, op cit, p. 46.
[237] Johnson, Phillip, in Woodward, Appendix 2, op. cit., p.219-220.
[238] Ralph O. Muncaster, Dismantling Evolution, (harvest House,
2003), p. 113.
[239] Darwin, op cit, p. 184.
[240] Dawkins, Richard, ĹDarwin Triumphantĺ, A Devilĺs Chaplain, op.
cit., p.86.
[241] Dawkins, Climbing Mount Improbable, op. cit.
[242] Vogel, Steven, Catĺs Paws and Catapults (Penguin, 1998),
p.23.
[243] Gould, Stephen Jay, ĹIs a new and general theory of evolution
emerging?ĺ, Paleobiology, Vol. 6 (1), January 1980, p.127.
[244] Gould, Stephen Jay, ĹThe Return of the Hopeful Monster,ĺ Natural
History, June/July 1977, p. 22, 24.
[245] Dawkins, A Devilĺs Chaplain, op. cit., p.211-212.
[248] Dennet, Daniel, ĹThe Leibnizian Paradigmĺ, David L. Hull and
Michael Ruse (ed.'s), The Philosophy of Biology (Oxford, 1998), p. 49.
[249] Denton, op. cit., p. 228, 315.
[250] Dembski, William A., ĹEvolutionĺs Logic of Credulity: An unfettered
response to Allen Orrĺ at www.arn.org/docs/dembski/wd_logic_credulity.htm
[251] Dawkins, The Blind Watchmaker, op. cit., p.146, 179,
my italics.
[252] Stone, David H., in John F. Ashton, (ed.), On the Seventh Day,
op. cit., p.90.
[253] Johnson, Phillip, ĹDaniel Dennettĺs Dangerous Ideaĺ, in Objection
Sustained (Downers Grove: IVP, 1998), p. 62.
[254] Denton, Natureĺs Destiny, op. cit., p.356.
[255] Harold, F., The
Way of the Cell: Molecules, Organisms and the Order of Life (New York: Oxford University Press, 2001).
[256] Denton, op. cit., p.331.
[257] Johnson, Phillip, ĹWhat is Darwinism?ĺ, in Objection Sustained,
(Downers Grove: IVP, 1998), p. 23.
[258] Dawkins, A Devilĺs Chaplain, op. cit., p.212.
[259] Wolf, Jakob, ĹTwo Kinds of Causality ľ Philosophical Reflections on
Darwinĺs Black Boxĺ at www.iscid.org/ubb/ultimatebb.php?ubb=get_topic;f=10;t=000040
[261] Dembski, No Free Lunch, op. cit., p.290.
[262] Darwin, Charles, Origin of Species, (1872), 6th edition,
(New York University Press, 1988), p. 154.
[263] Dawkins, Richard, ĹUniversal Darwinismĺ, in Hull and Ruse (ed.'s), The
Philosophy of Biology, op. cit., p.29.
[264] Dawkins, The Blind Watchmaker, op. cit., p.91.
[266] Piggliucci, ĹDesign Yes, Intelligent Noĺ, Darwin, Design,
And Public Education, p. 467.
[271] cf. Dembski,
William A., No Free Lunch (Oxford: Rowman & Littlefield, 2002),
p.279-289.
[272] Smart, John
A., ĹOn the Application of Irreducible Complexityĺ at www.iscid.org/papers/Smart_ApplicationOfIC_060503.pdf
[273] Behe, Michael
J., Dembski, Michael A. and Meyer, Stephen C., Science and Evidence for
Design in the Universe (San Francisco: Ignatius, 1999), 13-14.
[274] Dennet, Daniel
C., Darwinĺs Dangerous Idea (London: Penguin, 1995), p.318.
[276] Taylor, op cit, p. 118.
[278] Behe, Darwinĺs Black Box, op. cit., p.39.
[279] Gene, Mike, ĹIrreducible Complexity and Darwinian Pathwaysĺ at www.arn.org/docs/behe/mb_mg1darwinianpathways.htm
[280] Darwin, Origin of Species, op. cit., p 184.
[281] Dawkins, ĹDarwin Triumphantĺ, A Devilĺs Chaplain, op.
cit., p.86.
[282] Behe, op. cit., p. 40.
[285] Koons, op
cit, p. 14.
[286] Darwin, Charles, Origin of Species, (1872), 6th edition,
(New York University Press, 1988), p. 154.
[289] Behe, Michael J., ĹMolecular Machines: Experimental Support for the
Design Inferenceĺ at www.arn.org/docs/behe/mb_mm92496.htm
[290] Behe, Darwinĺs Black Box, op. cit., p.194.
[291] ĹSelf-Assembly of Bacterial Flagellaĺ at
www.aip.org/mgr/png/2002/174.htm
[292] Dembski, William A., ĹReinstating Design Within Scienceĺ, in Jay
Wesley Richards (ed.), Unapologetic Apologetics (Downers Grove: IVP,
2001), p.253.
[293] Behe, op. cit., p.72.
[294] Macnab, Robert M., CRC Critical Reviews in Biochemistry,
Vol. 5, Dec. 1978, p.291-341, quoted at
www.parentcompany.com/design_kit/dek1d.htm
[295] ĹHereĺs the recipe for making a just-so story . . . survey the
biological world for structures/functions. Find those that seem useful for
coming up with a precursor to the system in question and patch them together
without much regard for biochemical and/or genetic details. Place the patchwork
in an imaginary creature from the distant past that has conveniently gone
extinct. Invoke a vague selective pressure that selects for the patchwork and
then imagine it is plastic and amenable to further selective modification that
just happens to arrive at the system in question.ĺ ľ Julie Thomas, quoted by
Mike Gene, ĹEvolving the Bacterial Flagellum Through Cooptionĺ at
www.idthink.net/biot/flag1/
[296] Dennet, Darwinĺs Dangerous Idea, op. cit., p.521.
[297] TTSS secrete proteins to establish symbiotic relationships with
eukaryotic cells.
[298] Dembski, William A., ĹThe Bacterial Flagellum: Still Spinning Just
Fineĺ at www.designinference.com/documents/2003.02.Miller_Response.htm
[301] Orr, H. Allen, Boston Review.
[302] Thornhill, R. H. and Ussery, D. W., ĹA classification of possible
routes of Darwinian evolutionĺ, Journal of Theoretical Biology (2000)
203, p. 111-116.
[303] Minnich, Scott, in the video Unlocking the Mystery of Life.
[305] Gene, Mike, ĹIrreducible Complexity and Darwinian Pathwaysĺ at www.arn.org/docs/behe/mb_mg1darwinianpathways.htm
[306] Smart, ĹOn the Application of Irreducible Complexityĺ, op. cit.
[311] cf. www.iscid.org/pcid.php
[312] Bracht, J. R., ĹThe Bacterial Flagellum: A Response to Ursula
Goodenoughĺ at www.iscid.org/papers/Bracht_GoodenoughResponse_021203.pdf
[314] Dembski, William A., ĹGauging Intelligent Designĺs Successĺ at www.designinference.com/documents/2003.11.Gauging_IDs_Success.pdf
[315] Dembski, ĹThe Bacterial Flagellum: Still Spinning Just Fineĺ, op.
cit.
[316] Dembski, William
A., ĹIrreducible Complexity Revisitedĺ
[329] Dembski, William A., Intelligent Design (Downers Grove: IVP,
1999), p.149.
[330] Dembski, ĹIrreducible Complexity Revisitedĺ, op cit.
[331] Dembski, William A., The Intelligent Design Revolution (Downers Grove: IVP, 2004), p. 112-113.
[335] Behe, op cit, p. 40.
[336] Dembski, ĹGauging Intelligent Designĺs Successĺ, op. cit. cf. ĹThree Frequently Asked Questions About Intelligent Designĺ, op. cit. and
Axe, ĹExtreme Functional Sensitivity to Conservative Amino Acid Changes on
Enzyme Exteriorsĺ, op. cit.
[337] Darwin, Charles, Origin of Species, (1872), 6th edition,
(New York University Press, 1988), p. 154.
[339] Reppert, Victor, C.S. Lewisĺs Dangerous Idea, (IVP, 2003),
p. 116.
[341] Dubay, The Evidential Power of Beauty, op cit, p.
322.
[342] Crews, Frederick, ĹSaving us from Darwinĺ, New York Review of
Books.
[344] cf. ARN guide to Evolution, op cit.
[345] The fossil record does not reveal a gradual development from simple
to more complex species with numerous intermittent forms as Darwin predicted: Ĺthe record reveals the sudden appearance at differing times of
information-rich organisms within a hierarchical diversity of species with apparently
no precursors.ĺ ľ Francis J. Beckwith, Law, Darwinism, and Public Education:
The Establishment Clause and the Challenge of Intelligent Design, (Rowman
& Littlefield, 2003), p. 118.
[347] Dembski, op cit, p. 231.
[351] Dawkins, ĹDarwin Triumphantĺ, A Devilĺs Chaplin, op cit,
p. 81.
[353] ibid, p. 79, 81 & 84, my italics.
[355] This is what ID theorists tend to mean by ĹDarwinismĺ: belief in
Ĺthe fact of evolution itselfĺ wedded to a philosophical commitment to the
explanatory sufficiency of natural processes (usually expressed as the
multi-faceted theory that life arose millions of years ago by purely natural
processes and then diversified from a common ancestor over millions of years
through the undirected but non-random survival of small random hereditary
changes).
[356] W.R. Thompson, introduction to Charles Darwin, The Origin of
Species by Means of Natural Selection, (New York: Dutton, 1967), pp. xxii.
[357] Lee Strobel, The Case for Faith, (Grand Rapids: Zondervan,
2000), p. 90.
[358] Gordon Graham, Genes: A Philosophical Inquiry, (London: Routledge, 2002), p ix.
[360] David Stove, quoted by Graham, ibid, p 31.
[363] Norman L. Geisler and Joseph Holden, Living Loud: Defending Your
Faith, (Nashville, Tennessee: Broadman and Holman Publishers, 2002), p. 55.
[364] William Lane Craig, ĹTough Questions About Scienceĺ, in Ravi
Zacharias & Norman L. Geisler (ed.ĺs), Who Made God?, (Grand Rapids: Zondervan, 2003), p. 70.
[366] Dawkins, A Devilĺs Chaplain, op cit, p. 220.
[367] Jonathan Wells, ĹThe Intelligent Design Movement, Evangelical
Scientists, and the Future of Biology,ĺ in Darwinism Defeated?, op
cit, p. 137.
[368] Cliff Marsh, ĹEvolution: a theory in terminal decline?ĺ, The
Plain Truth, September-November 2002, p. 20.
[369] Michael Denton, quoted from his professional resume, cf. Woodward, op
cit, p. 57.
[374] Paul Davies, Superforce, (Touchstone Books, 1985), p. 243.
[375] Paul Davies, The Cosmic Blueprint, (London: Harper Collins,
1989), p. 203.
[376] Paul Davies, The Fifth Miracle: The Search for the Origin of
Life, (London: Penguin Press, 1998), p. xvi.
[381] Wiker, ĹDoes Science Point to God?ĺ, op cit.
[382] Denis Alexander, Rebuilding the Matrix, (Oxford: Lion,
2001), p. 21.
[383] Phillip E. Johnson, The Right Questions, (Downers Grove:
IVP, 2002), p. 80.
[384] Woodward, op cit, p. 253.
[385] Charles Darwin, quoted in N. C. Gillespie, Charles Darwin and
the Problem of Creation, (Chicago: University of Chicago, 1979), p. 87.
[386] Charles Darwin, The Origin of Species, (Ware: Wordsworth
Editions Limited, 1998), p. 363.
[387] Dembski, ĹThree Frequently Asked Questions About Intelligent
Designĺ, op cit.
[388] Dembski, Intelligent Design, op cit, p. 113.
[389] Benjamin Wiker, Moral Darwinism, (Downers Grove: IVP, 2002),
p. 295.
[391] Dembski, No Free Lunch, op cit, p. 246.
[392] Dembski, ĹOn the Very Possibility of Intelligent Designĺ, The
Creation Hypothesis, op cit, p. 132.
[393] Dembski, Intelligent Design, op cit, p. 113.
[395] Dembski, ĹEvolutionĺs Logic of Credulity: An Unfettered Response to
Allen Orrĺ, op cit.
[397] Wiker, ĹDoes Science Point to God?ĺ, op cit.
[399] Dawkins, A Devilĺs Chaplin, op cit, p. 58.
[400] Theodosius Dobzhansky, quoted by Wells, op cit, p. 329.
[401] Wells, ibid, p. 329.
[402] Dembski, Intelligent Design, op cit, p. 255.
[403] Wiker, op cit, p. 3.
[406] Dembski, op cit, p. 256-257.
[407] Woodward, op cit, p. 210.
[413] Dembski, Intelligent Design, op cit, p. 255-256.
[415] cf. Dembski, ĹBecoming a Disciplined Scienceĺ, op cit.
[417] Paul Davies, quoted by Witham, By Design, (San Francisco:
Encounter Books, 2003), p. 149.
[419] Dembski, The Design Revolution, op cit, p. 305.
[420] Huston Smith, quoted on the front cover of On the Seventh Day,
op cit, front cover quote.
)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}