Second Thoughts about Peppered Moths
This classical story of evolution by natural selection needs revising
Jonathan Wells, Ph.D.
Department of Molecular Cell Biology
University of California
Berkeley, California, USA
Every student of biological evolution learns about peppered moths. The dramatic increase in dark forms of this species during the industrial revolution, and experiments pointing to differential bird predation as the cause, have become the classical story of evolution by natural selection. The same careful scientific approach which established the classical story in the first place, however, has now revealed major flaws in it. It is time to take another look.
The peppered moth, Biston betularia, comes in various shades of gray. One hundred and fifty years ago, the species consisted almost entirely of "typical" forms, with predominantly light gray scales interspersed with black (hence the name, "peppered"). In 1848, a coal-black "melanic" form was collected near Manchester, England, and by 1950 melanic forms made up more than 90% of the peppered moths in that area. A similar change was reported in many other species of moths, as well as in ladybird beetles, spiders, and even some birds. Since the phenomenon was observed not only around Manchester but also near other industrial centers such as Birmingham and Liverpool, it became known as "industrial melanism."
The Classical Story
Before 1950, the cause of industrial melanism was largely a matter of speculation. According to Tutt (1896), the cause was cryptic coloration and differential predation. Tutt theorized that in unpolluted woodlands, typicals are well camouflaged against the light-colored lichens that grow on tree trunks; but in woodlands where industrial pollution has killed the lichens and exposed the dark brown tree trunks, melanics are better camouflaged (Figure 1). Since predatory birds are more likely to eat the most conspicuous moths, melanic forms would increase as a result of natural selection. Tutt's theory, though plausible, was not widely accepted at the time because entomologists and ornithologists had no evidence that birds were major predators of camouflaged day-resting moths.
 |
 |
 |
| All Three Images © Laurence M. Cook |
|
|
| Figure 1. Peppered moths resting on three different tree trunks. (top) Two moths (one typical and one melanic) resting on the dark bark of an oak tree near the industrial city of Liverpool, U.K. (middle) The same two moths on a nearby beech tree covered by a combination of green algae and lichen. (bottom) Typical and melanic moths resting on light-colored lichen on an oak tree in rural Wales. Note the striking differences in camouflage efficiency (Bishop and Cook 1975; used by permission). |
|
Harrison (1920) rejected Tutt's natural selection theory, and proposed instead that melanism was induced directly by airborne industrial pollutants. Although he did not work on Biston betularia, Harrison reported that melanism could be produced in several other moth species if their larvae were fed on leaves contaminated with metallic salts. Critics pointed out, however, that some of the species Harrison used did not exhibit industrial melanism in the wild. Ford (1937) rejected Harrison's induction theory in favor of natural selection; but he also rejected Tutt's theory that the selective agent was bird predation, and maintained that melanic forms in polluted environments enjoyed a physiological advantage unrelated to their color (reviewed in Majerus 1998).
Kettlewell's experiments. In the 1950's, British entomologist Bernard Kettlewell set out to resolve the issue empirically. Like Tutt, Kettlewell believed that industrial melanism was due to cryptic coloration and selective predation, and he used three experimental approaches to test the theory. First, he estimated the moths' camouflage efficiency on various backgrounds, as judged by the human eye. Second, he directly observed bird predation through binoculars. Third, he marked and released larger numbers of moths, then recaptured some and compared the pre-release to post-recapture proportions.
Kettlewell's first experiment was conducted in an aviary containing a pair of nesting birds and their young. He released peppered moths into the aviary, and watched through binoculars as they settled onto resting sites and were subsequently eaten by the birds. Kettlewell (1955) thus established that birds do, in fact, prey on resting peppered moths.
The second experiment consisted of marking several hundred peppered moths, including typicals as well as melanics, and releasing them into a polluted woodland near Birmingham, England. Kettlewell watched through binoculars as the moths settled on nearby trees; he observed that melanics were much less conspicuous than typicals, as judged by the human eye, and that birds took conspicuous moths more readily than inconspicuous ones. That night he set out traps to recapture as many as he could. Of 447 released melanics he recaptured 123, while of 137 released typicals he recaptured only 18. In other words, he recaptured 27.5% of the melanics, but only 13.0% of the typicals, suggesting that a much higher proportion of melanics had survived predation. Kettlewell concluded that "birds act as selective agents, as postulated by evolutionary theory" (Kettlewell 1955, p. 342).
In Kettlewell's third experiment, the same procedure was repeated in an unpolluted woodland in Dorset, England. Once again he released moths, scored them for crypsis, and watched for selective predation. Ethologist Niko Tinbergen accompanied him and made movies of birds picking the moths off tree trunks. As expected, melanic moths were much more conspicuous than typicals on the lichen-covered Dorset trees, and thus more readily taken by predatory birds. Kettlewell also repeated his mark-release-recapture experiment, and the results were the opposite of those obtained in the Birmingham experiment. He recaptured 62 of 496 released typicals (12.5%), but only 30 of 473 released melanics (6.3%), so the 2:1 recapture ratio which he had found in Birmingham was completely reversed. Kettlewell concluded that typicals enjoyed a selective advantage in Dorset because their superior camouflage improved their chances of surviving bird predation. He then returned to Birmingham so Tinbergen could make movies of selective predation in a polluted woodland to complement the movies he had made in an unpolluted one (Kettlewell 1956).
Further support for Kettlewell's theory. Other biologists conducted experiments with peppered moths on tree trunks to test Kettlewell's theory that industrial melanism was due to cryptic coloration and selective predation (e.g., Clarke and Sheppard 1966, Bishop 1972, Lees and Creed 1975, Bishop and Cook 1975, Steward 1977b, Murray et al. 1980). Their conclusions generally agreed with Kettlewell's.
Following the passage of anti-pollution legislation in the 1950's, industrial melanism began to decline. The percentage of melanics on the Wirral Peninsula, west of Liverpool, dropped from 93% in 1959 to 90% in 1962 (Clarke and Sheppard 1966, Kettlewell 1973). A decade later, Bishop and Cook (1975) reported that the reversal of industrial melanism was well under way. A reversal was also noted in other urban areas as air quality improved (reviewed in Lees 1981). By 1984, the percentage of melanics on the Wirral Peninsula had decreased to 61% (Clarke et al. 1985), and in 1996 the percentage was only 8.5% (Grant et al. 1998). Since pollution-control legislation would be expected to restore the typicals' cryptic advantage by enabling lichens to return to the trees, the decline of industrial melanism was consistent with cryptic coloration and selective predation, and was thus widely regarded as further confirmation of Kettlewell's theory.
The textbook example of evolution in action. Kettlewell described industrial melanism in moths as "the most striking evolutionary change ever actually witnessed in any organism" (Kettlewell 1955, p. 323). Two decades later, British geneticist P. M. Sheppard called it "the most spectacular evolutionary change ever witnessed and recorded by man, with the possible exception of some examples of pesticide resistance" (Sheppard 1975, p. 70), and famed population geneticist Sewall Wright called it "the clearest case in which a conspicuous evolutionary process has actually been observed" (Wright 1978, p. 186).
Industrial melanism in peppered moths thereby became "the textbook example of natural selection in action" (Jones 1982, p. 109). According to Majerus, "everyone knows the basic peppered moth story, because it is in all the textbooks" (Majerus 1998, p. 97). One recent evolution textbook calls it "a classic example of natural selection" which is "perhaps the best-known story in evolutionary biology," though the story is "now known to be more complex" than originally thought (Ridley 1996, pp. 103-109).
According to Majerus, however, evidence gathered in the 40 years since Kettlewell's work shows that "the basic peppered moth story is wrong, inaccurate, or incomplete, with respect to most of the story's component parts" (Majerus 1998, p. 116). What follows is a review of some of the flaws that have been discovered in the classical story.
Problems with the Classical Story
When biologists looked beyond Birmingham and Dorset, where Kettlewell had conducted his experiments, they found discrepancies between Kettlewell's theory and the actual geographical distribution of melanic moths. For example, if melanic moths in polluted woodlands enjoyed as much of a selective advantage as Kettlewell's experiments seemed to indicate, then they should have completely replaced typicals in heavily polluted areas such as Manchester (Bishop and Cook 1980, Mani 1990). This never happened, however, indicating that factors other than selective predation must be affecting melanic frequencies. Some investigators attributed the discrepancy to heterozygote advantage (Clarke and Sheppard 1966, Lees and Creed 1975), but it has since been established that there is no evidence for this (Creed et al. 1980, Lees 1981, Mani 1982, Cook et al. 1986).
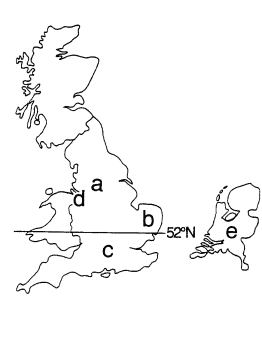 |
Figure 2. Problems with the classical story: The U.K. and The Netherlands. (a) Despite theoretical predictions, the proportion of melanics around heavily-polluted Manchester never reached 100% (Bishop and Cook 1980, Mani 1990). (b) The proportion of melanics in East Anglia reached 80% despite the absence of any apparent pollution (Lees and Creed 1975); after the introduction of pollution control legislation, typicals became predominant before lichens returned to the trees (Grant and Howlett 1988). (c) South of latitude 52*N, the relatively poor correlation of melanism with sulfur dioxide concentration suggested that non-industrial factors were of greater importance than selective predation (Steward 1977a,b); after the introduction of pollution control legislation, the proportion of melanics decreased in the north, as expected, but increased in the south (Bishop and Cook 1980, Jones 1982). (d) The frequency of typicals on the Wirral Peninsula increased dramatically before the return of lichens to tree trunks (Clarke et al. 1985, Grant et al. 1998). (e) The decline of melanism in The Netherlands has been accompanied by an increase not only in typicals, but also in an intermediate form almost as dark as melanics. (Brakefield 1990). |
|
Some other distribution features were inconsistent with Kettlewell's explanation, as well. In rural Wales, the frequency of melanics was higher than expected, leading Bishop to conclude that "as yet unknown factors" were involved (Bishop 1972, p. 240). In rural East Anglia, where there was little industrial pollution and typicals seemed better camouflaged, melanics reached a frequency of 80%, prompting Lees and Creed to write: "We conclude therefore that either the predation experiments and tests of conspicuousness to humans are misleading, or some factors or factors in addition to selective predation are responsible for maintaining the high melanic frequencies" (Lees and Creed 1975, pp. 75-76).
On the other hand, melanics in south Wales were better camouflaged than typicals, yet comprised only about 20% of the population (Steward 1977a,b). Compiling data from 165 separate sites in Britain, Steward found a correlation between melanism and the concentration of sulfur dioxide (an airborne pollutant) north of latitude 52*N (Figure 2), but concluded that "in the south of Britain non-industrial factors may be of greater importance" than selective predation. Steward cautioned that "it may not be possible to generalize from the results for one area, to explain geographic variation over the rest of Britain" (Steward 1977a, pp. 239, 242).
After the passage of anti-pollution legislation, the proportion of melanics decreased north of London (as expected), but inexplicably increased to the south (Bishop and Cook 1980; Jones 1982). In The Netherlands, the decline of melanism took another twist. As air pollution declined, not only did the frequency of typicals increase, but also the frequency of an intermediate form which was almost as dark as melanics, suggesting a more complex change than was seen in Britain (Brakefield 1990).
Various theoretical models have been proposed to account for the discrepancies. Some include the effects of gene flow due to migration, though according to Jones "gene flow alone cannot explain... why melanics are so common in some unpolluted parts of Britain" (Jones 1982, p. 109). Mani (1990), like Steward (1977a), obtained a good fit between melanism and sulfur dioxide concentration, but cautioned that "such a correlation does not define causal connection. It only says that SO2 concentration can be used as an approximate measure of the level of pollution that affects the morphs differentially in some unknown way" (Mani 1990, p. 368; emphasis in original). Whatever the actual causes may be, Berry concluded, "it is clear that melanic peppered moth frequencies are determined by much more than differential visual predation by birds" (Berry 1990, p. 312).
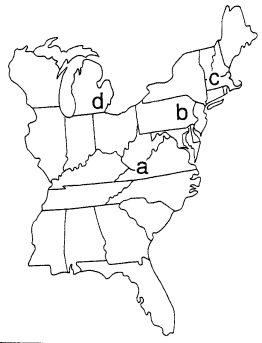
|
Figure 3. Problems with the classical story: The U. S. (a) In the 1970’s, the frequency of melanics in an unspoiled forest in southwestern Virginia was about double the frequency in polluted Blacksburg 18 km away; neither lichen cover nor gene flow could explain the difference (West 1977). (b) From 1971 to 1978, melanics remained at about 52% in a low-pollution area in central eastern Pennsylvania (Manley 1981). (c) Melanics occurred at low frequencies in western and central Massachusetts even though trees were neither devoid of lichens nor blackened by soot (Sargent 1974, Treat 1979). (d) Between the 1960’s and 1990’s, melanics in southeastern Michigan increased and decreased in parallel with those in England, but without any perceptible changes in lichen cover (Grant et al. 1996). |
|
In the United States, the first melanic peppered moth was reported in Philadelphia in 1906, and melanism increased until by 1959 it was over 90% in some areas (Owen 1962). Yet the geographical distribution did not fit the classical story any better than it did in the U.K. (Figure 3). In the 1970’s, the frequency of melanics in a seemingly unspoiled forest in southwestern Virginia was more than double the frequency at polluted Blacksburg, 18 km away. The difference was not related to lichen cover, and could not be explained by pollution levels or gene flow (West 1977). In a low-pollution area in central eastern Pennsylvania, melanics remained at about 52% from 1971 to 1978 (Manley 1981), and in western and central Massachusetts melanics persisted at low frequencies even though the trees there were neither devoid of lichens nor blackened by soot (Sargent 1974, Treat 1979). In southeastern Michigan, the frequency of melanics was over 90% in the early 1960's, then dropped to less than 20% by 1995 (Grant et al. 1995), thus paralleling the decline of melanism in the U.K. Yet the decline in Michigan "occurred in the absence of perceptible changes in local lichen floras" (Grant et al. 1996, p. 351).
Recently, Grant and his colleagues reported a good correlation between sulfur dioxide levels and melanic frequencies in southwestern Virginia, central eastern Pennsylvania, and southeastern Michigan. In fact, the decline of melanism in both the U.S. and the U.K. appears to be "correlated primarily with reductions in atmospheric sulfur dioxide" (Grant et al. 1998, p. 465). The lack of correlation between between melanic frequencies and lichen cover, however, is a serious problem for the theory that industrial melanism is due to cryptic coloration and selective predation. This aspect of the story warrants a closer look.
Is there a correlation between melanism and lichen cover? In the 1970's, Kettlewell noted that melanism began declining on the Wirral Peninsula before lichens returned to the trees (Kettlewell 1973). Lees and his colleagues surveyed melanism in peppered moths at 104 sites throughout Britain, and found a lack of correlation with lichen cover which they considered "surprising in view of the results of Kettlewell's selection experiments" (Lees et al. 1973). As noted above, American biologists during the same period reported that melanism was also unrelated to lichen cover in the U.S. (Sargent 1974, West 1977, Treat 1979).
The discrepancy has persisted. In the early 1980's, Clarke and his colleagues found "a reasonable correlation" in the U.K. between the decline in melanism and decrease in sulfur dioxide pollution, but were surprised to note "that throughout this time the appearance of the trees in Wirral does not seem to have changed appreciably" (Clarke et al. 1985, p. 198). According to Grant and Howlett, if the rise of industrial melanism was originally due to the demise of lichens on trees, then "the prediction is that lichens should precede the recovery of the typical morph as the common form. That is, the hiding places should recover before the hidden. But, this is clearly not the case in at least two regions where the recovery of typicals has been especially well documented in the virtual absence of these lichens: on the Wirral..., and in East Anglia" (Grant and Howlett 1988, pp. 230-231). Ten years later, the situation on the Wirral Peninsula remains the same; Grant and his colleagues note that "even now lichen coverage on trees in that region is sparse, yet the typicals exceed 93%" (Grant et al. 1998, p. 466).
The decline of melanism in the U.S. without perceptible changes in the lichen cover prompted Grant and his colleagues to conclude that "the role of lichens has been inappropriately emphasized in chronicles about the evolution of melanism in peppered moths" (Grant et al. 1996, p. 351). Sargent and his colleagues note that "the recent declining frequency of melanism in B. betularia in North America, where the hypothesis of a cryptic advantage of melanism never seemed applicable," is "perplexing" in view of the classical story (Sargent et al. 1998, pp. 316-317). So melanism declined in the U.K. before lichens returned to the trees. In the U.S., melanism occurred in the presence of lichens and declined without any perceptible change in them. Clearly, the rise and fall of industrial melanism did not depend on the presence or absence of lichens. Yet their presence or absence seemed highly significant in Kettlewell's experiments. Why the discrepancy?
The normal resting places of peppered moths. In most of Kettlewell's experiments, moths were released in the morning and observed during the day. Recaptures were made at night. In only one experiment (June 18, 1955) did Kettlewell release moths before sunrise; he abandoned this approach because of practical difficulties such as having to warm the cold moths beforehand on the engine of his car (Kettlewell 1973).
But peppered moths are night-fliers, and normally find resting places on trees before dawn. Artificial daytime releases might have disoriented the moths, causing them to fly immediately to exposed tree trunks where they became unnaturally easy targets for predatory birds. Regarding his release methods, Kettlewell wrote: "To the obvious criticism that the releases were not free to take up their own choice of resting site for the first day, I must answer that there were no other alternative backgrounds available for an insect that has to spend its days on trunks and boughs in this wood. I admit that, under their own choice, many would have taken up position higher in the trees, and... in so doing they would have avoided concentrations such as I produced.... [O]thers have shown the importance to cryptic insects of avoiding too high a density level, but this is no argument against the findings for the relative advantages" of the light and dark forms. Kettlewell granted, however, "that, under natural conditions, predation, though selective, might take place at a lower tempo" (Kettlewell 1955, p. 340; emphasis in original). In other words, Kettlewell assumed (1) that the main defect of his release method was an unnaturally high density of moths, affecting merely the tempo of predation; and (2) that he could disregard the observation that many moths would have preferred to take up positions higher in the trees.
Before the 1980's most investigators shared Kettlewell's second assumption, and many of them found it convenient to conduct predation experiments using dead specimens glued or pinned to tree trunks (e.g., Clarke and Sheppard 1966, Bishop 1972, Lees and Creed 1975, Bishop and Cook 1975, Steward 1977b, Murray et al. 1980). Kettlewell himself considered this a bad idea (Kettlewell 1973), and some biologists who used dead moths suspected that the technique was unsatisfactory. For example, Bishop and Cook conducted predation experiments using dead moths glued to trees; but they noted discrepancies in their results which "may indicate that we are not correctly assessing the true nature of the resting sites of living moths when we are conducting experiments with dead ones" (Bishop and Cook 1975, p. 9).
Figure 1, which is taken from Bishop and Cook's 1975 article, shows live moths rather than dead ones, but because peppered moths are quite torpid during the day it was easy to place them manually on the desired backgrounds (L. M. Cook, 1998, personal communication. University of Manchester, Manchester, U.K.). Indeed, close inspection of Figure 1 reveals that the pattern of gray-and-black scales in the typical form, and the pattern of wing veins in the melanic form, are identical in the top two panels. Clearly, the same two moths were placed on two different backgrounds. Most textbook photographs of peppered moths similarly show specimens which have been manually placed on tree trunks (Sargent et al. 1998).
Since 1980, however, evidence has accumulated showing that peppered moths do not normally rest on tree trunks. Using caged moths, Mikkola observed that "the normal resting place of the Peppered Moth is beneath small, more or less horizontal branches (but not on narrow twigs), probably high up in the canopies, and the species probably only exceptionally rests on tree trunks." He noted that "night-active moths, released in an illumination bright enough for the human eye, may well choose their resting sites as soon as possible and most probably atypically." Thus "the results of Kettlewell (1955, 1956) fail to demonstrate the qualitative predation of the morphs of the Peppered Moth by birds or other predators in natural conditions" (Mikkola 1984, pp. 416-418).
Mikkola used caged moths, but data on wild moths support his conclusion. In twenty-five years of field work, Clarke and his colleagues found only one peppered moth on a tree trunk, and admitted that they knew primarily "where the moths do not spend the day" (Clarke et al. 1985, p. 197; emphasis in original). When Howlett and Majerus studied the natural resting sites of peppered moths in various parts of England, they found that Mikkola's observations on caged moths were valid for wild moths, as well. They concluded: "...it seems certain that most B. betularia rest where they are hidden.... [and] that exposed areas of tree trunks are not an important resting site for any form of B. betularia" (Howlett and Majerus 1987, p. 40). In a separate study, Liebert and Brakefield confirmed Mikkola's observations that "the species rests predominantly on branches.... Many moths will rest underneath, or on the side of, narrow branches in the canopy" (Liebert and Brakefield 1987, p. 129).
In a recent book on melanism, Majerus criticizes the "artificiality" of much previous work in this area, noting that "in most predation experiments peppered moths have been positioned on vertical tree trunks, despite the fact that they rarely chose such surfaces to rest upon in the wild" (Majerus 1998, p. 116). If peppered moths normally rest under horizontal branches in the upper canopy, then observations of differential bird predation on moths which are placed on tree trunks probably have little relevance to their survival in the wild. It appears that the classical example of natural selection is actually be an example of unnatural selection!
Conclusions
Bernard Kettlewell was a good scientist. Even now, almost half a century after his initial experiments, Kettlewell's scientific papers make exciting reading. But science doesn't stop with initial experiments. Kettlewell's successors, who were also good scientists, found unexpected problems with his conclusion that industrial melanism is due to cryptic coloration and selective predation.
The geographic distribution of melanic peppered moths did not fit the theory: the frequency of melanics was not as high as it should have been in some places, and higher than it should have been in others. Furthermore, melanism is not correlated with lichen cover; in the U.K., it declined before lichens returned to the trees, while in the U.S., it occurred despite the presence of lichens and declined without any perceptible changes in lichen cover. Finally, peppered moths do not normally rest on tree trunks: instead, they normally rest under horizontal branches high in the canopy, not where Kettlewell and his successors had carried out their experiments on selective predation.
These findings do not entirely rule out a role for cryptic coloration and selective predation in industrial melanism, but at the very least they deprive Kettlewell's explanation of empirical support. As one recent review concludes, "there is little persuasive evidence, in the form of rigorous and replicated observations and experiments, to support this explanation at the present time" (Sargent et al. 1998, p. 318).
In at least one insect (the ladybird beetle, Adalia bipunctata), industrial melanism has nothing to do with cryptic coloration and selective predation. Ladybird beetles are extremely distasteful to most birds, and exhibit warning coloration rather than cryptic coloration; there is no evidence that they are significantly affected by selective predation (Creed 1966, Ford 1975). Industrial melanism in ladybird beetles has been attributed to non-visual selection on the ability of melanic and non-melanic forms to absorb solar radiation -- a phenomenon known as "thermal melanism" (Brakefield 1985).
This does not mean that the same explanation applies to peppered moths, but it clearly indicates that cryptic coloration and selective predation are not the only possible explanations for industrial melanism. Some biologists continue to believe, like Harrison (1920), that melanism might be directly induced by environmental factors (reviewed in Sargent et al. 1998). Most biologists, however, believe that natural selection is responsible, though no one knows what traits are being selected or what factors in the environment are doing the selecting.
The very prominence of the peppered moth story in the teaching of evolution requires that it be scrupulously accurate. According to Grant and Howlett, "as Biston betularia has served as a paradigm of evolution, it demands the closest possible scrutiny" (Grant and Howlett 1988, p. 231). Yet this classical story of evolution by natural selection, as it continues to be retold in many textbooks, is seriously flawed. In particular, the illustrations which typically accompany the story (like the photographs in Figure 1) mislead students by portraying peppered moths on tree trunks where they do not normally rest. Unknown to Kettlewell, his experiments had less to do with natural selection than with unnatural selection, and the true causes of industrial melanism in peppered moths remain largely unknown.
The classical story, elegant and appealing though it may be, should no longer be presented as a textbook example of evolution in action. If the purpose of science education is to teach students how to do good science, then instead of re-telling the classical story textbooks would do better to focus on how science revealed its flaws.
Acknowledgments
The author thanks Laurence M. Cook for helpful comments and for permission to use his 1975 photograph of peppered moths, and thanks Bruce Grant for supplying a preprint of his 1998 article in Journal of Heredity. The author also thanks Lucy Wells and Roberta Bidinger for their assistance with the manuscript. Research for this article was funded by a grant from The Discovery Institute, Seattle, WA.
References
Berry RJ. 1990. Industrial melanism and peppered moths (Biston betularia (L.). Biological Journal of the Linnean Society 39: 301-322.
Bishop JA. 1972. An experimental study of the cline of industrial melanism in Biston betularia (L.) (Lepidoptera) between urban Liverpool and rural North Wales. Journal of Animal Ecology 41: 209-243.
Bishop JA, Cook LM. 1975. Moths, melanism and clean air. Scientific American 232(1): 90-99.
Bishop JA, Cook LM. 1980. Industrial melanism and the urban environment. Advances in Ecological Research 11: 373-404.
Brakefield PM. 1985. Polymorphic Muellerian mimicry and interactions with thermal melanism in ladybirds and a soldier beetle: a hypothesis. Biological Journal of the Linnean Society 26: 243-267.
Brakefield PM. 1990. A decline of melanism in the peppered moth Biston betularia in The Netherlands. Biological Journal of the Linnean Society 39: 327-334.
Clarke CA, Sheppard PM. 1966. A local survey of the distribution of industrial melanic forms in the moth Biston betularia and estimates of the selective values of these in an industrial environment. Proceedings of the Royal Society of London B 165: 424-439.
Clarke CA, Mani GS, Wynne G. 1985. Evolution in reverse: clean air and the peppered moth. Biological Journal of the Linnean Society 26: 189- 199.
Cook LM, Mani GS, Varley ME. 1986. Postindustrial melanism in the peppered moth. Science 231: 611-613.
Creed ER. 1966. Geographic variation in the two-spot ladybird in England and Wales. Heredity 21: 57-72.
Creed ER, Lees DR, Bulmer MG. 1980. Pre-adult viability differences of melanic Biston betularia (L.) (Lepidoptera). Biological Journal of the Linnean Society 13: 251-262.
Ford EB. 1937. Problems of heredity in the Lepidoptera. Biological Reviews 12: 461-503.
Ford EB. 1975. Ecological Genetics. 4th ed. London: Chapman and Hall.
Grant BS, Howlett RJ. 1988. Background selection by the peppered moth (Biston betularia Linn.): individual differences. Biological Journal of the Linnean Society 33: 217-232.
Grant BS, Owen DF, Clarke CA. 1995. Decline of melanic moths. Nature 373: 565.
Grant BS, Owen DF, Clarke CA. 1996. Parallel rise and fall of melanic peppered moths in America and Britain. Journal of Heredity 87: 351- 357.
Grant BS, Cook AD, Clarke CA, Owen DF. 1998. Geographic and temporal variation in the incidence of melanism in peppered moth populations in America and Britain. Journal of Heredity 89: 465-471.
Harrison JWH. 1920. Genetical studies in the moths of the geometrid genus Oporabia (Oporinia) with a special consideration of melanism in the Lepidoptera. Journal of Genetics 9: 195-280.
Howlett RJ, Majerus MEN. 1987. The understanding of industrial melanism in the peppered moth (Biston betularia) (Lepidoptera: Geometridae). Biological Journal of the Linnean Society 30: 31-44.
Jones JS. 1982. More to melanism than meets the eye. Nature 300: 109-110.
Kettlewell HBD. 1955. Selection experiments on industrial melanism in the Lepidoptera. Heredity 9: 323-342.
Kettlewell HBD. 1956. Further selection experiments on industrial melanism in the Lepidoptera. Heredity 10: 287-301.
Kettlewell HBD. 1973. The Evolution of Melanism. Oxford: Clarendon Press.
Lees DR. 1981. Industrial melanism: genetic adaptation of animals to air pollution. Pages 129-176 in Bishop JA, Cook LM, eds. Genetic Consequences of Man-made Change. London: Academic Press.
Lees DR, Creed ER. 1975. Industrial melanism in Biston betularia: the role of selective predation. Journal of Animal Ecology 44: 67-83.
Lees DR, Creed ER, Duckett LG. 1973. Atmospheric pollution and industrial melanism. Heredity 30: 227-232.
Liebert TG, Brakefield PM. 1987. Behavioural studies on the peppered moth Biston betularia and a discussion of the role of pollution and lichens in industrial melanism. Biological Journal of the Linnean Society 31: 129- 150.
Majerus MEN. 1998. Melanism: Evolution in Action. Oxford: Oxford University Press.
Mani GS. 1982. A theoretical analysis of the morph frequency variation in the peppered moth over England and Wales. Biological Journal of the Linnean Society 17: 259-267.
Mani GS. 1990. Theoretical models of melanism in Biston betularia -- a review. Biological Journal of the Linnean Society 39: 355-371.
Manley TR. 1981. Frequencies of the melanic morph of Biston cognataria (Geometridae) in a low-pollution area of Pennsylvania from 1971 to 1978. Journal of the Lepidopterists' Society 35: 257-265.
Mikkola K. 1984. On the selective forces acting in the industrial melanism of Biston and Oligia moths (Lepidoptera: Geometridae and Noctuidae). Biological Journal of the Linnean Society 21: 409-421.
Murray ND, Bishop JA, MacNair MR. 1980. Melanism and predation by birds in the moths Biston betularia and Phigalia pilosauria. Proceedings of the Royal Society of London B 210: 277-283.
Owen DF. 1962. The evolution of melanism in six species of North American geometrid moths. Annals of the Entomological Society of America 55: 695-703.
Ridley M. 1996. Evolution. 2nd ed. Cambridge (MA): Blackwell Science.
Sargent TD. 1974. Melanism in moths of central Massachusetts (Noctuidae, Geometridae). Journal of the Lepidopterists' Society 28: 145-152.
Sargent TD, Millar CD, Lambert DM. 1998. The "classical" explanation of industrial melanism: assessing the evidence. Evolutionary Biology 30: 299-322.
Sheppard PM. 1975. Natural Selection and Heredity. 4th ed. London: Hutchinson University Library.
Steward RC. 1977a. Industrial and non-industrial melanism in the peppered moth, Biston betularia (L.). Ecological Entomology 2: 231-243.
Steward RC. 1977b. Melanism and selective predation in three species of moths. Journal of Animal Ecology 46: 483-496.
Treat AE. 1979. Biston cognataria (Geometridae): frequency of melanic males in Tyringham, Massachusetts, 1958-1977. Journal of the Lepidopterists' Society 33: 148-149.
Tutt JW. 1896. British Moths. London: George Routledge.
West DA. 1977. Melanism in Biston (Lepidoptera: Geometridae) in the rural central Appalachians. Heredity 39: 75-81.
Wright S. 1978. Evolution and the Genetics of Populations. Volume 4: Variability Within and Among Natural Populations. Chicago: The University of Chicago Press.
[Updated: April 6, 1999]
Copyright © 1999 by Jonathan Wells. All rights reserved. International copyright secured.
File Date: 4.10.99