
Malcolm S. Gordon, “The Concept of Monophyly: A Speculative Essay,” Biology and Philosophy 14 (1999):331-348.
The concept of common descent, or monophyly, is held by modern biologists as a “near ideological commitment” (p. 332), says Malcolm Gordon, professor of biology at UCLA. In this remarkable essay, Gordon looks at two episodes in the history of life—the origin of life itself and the origin of tetrapods–and argues that in both cases, the evidence favors multiple, or polyphyletic, origins. The standard concept of common ancestry is well supported only at the smaller scales of taxonomic differentiation, Gordon asserts, namely, “from populations to the levels of genera and families” (p. 333), although he adds that “there are no single, universal scenarios, even at these levels.”
The least convincing applications of monophyly, Gordon continues, “involve the macro-scale of evolutionary differentiation, the highest categories (kingdoms, phyla, classes)” (p. 333). His skepticism is “based upon discoveries in recent years flowing primarily from genome sequencing projects and other aspects of molecular genetics” (p. 334). The crucial question is this: “Do traceable genetic lines of descent exist that might ultimately permit characterization of the genomes of organisms basal to the clades for the highest categories?” Gordon responds: “The answer to this question increasingly appears to be no” (p. 334; emphasis in original). He lists seven major sets of phenomena and processes which may have scrambled irretrievably any phylogenetic signal in genomes (e.g., gene transfer, hybridization), and concludes: “These phenomena collectively make it appear nearly impossible that the highest categories of living organisms can be said to have had basal species” (p. 335).
Thus, “at the macro-scale, life appears to have had many origins. The base of the tree of life appears not to have been a single root, but was instead a network of inextricably intertwined multiple branches deriving from many, perhaps 100 or more, genetic sources. . . . The traditional version of the theory of common descent apparently does not apply to kingdoms as presently recognized. It probably does not apply to many, if not all phyla, and possibly also not to many classes within the phyla” (p. 335).
“Life, therefore, is probably polyphyletic in origin,” he concludes (p. 337).
Gordon’s discussion of tetrapods is equally interesting. He points out that most analyses of the origin of tetrapod begin by assuming common ancestry. Polyphyletic (multiple) origins are excluded, therefore, not on the basis of the evidence but of philosophy. As he explains, “the analyses were all done cladistically,” and hence “the underlying phylogenetic model in all cases was monophyletic. A single ‘main line’ of tetrapod evolution is assumed to have existed in all cases. Possible polyphyletic scenarios were methodologically and philosophically excluded as implausible” (p. 339).
The fossil record for tetrapods is, at any rate, “limited and partial.” Though “a few forms are now represented by well-preserved, near complete remains (notably Eusthenopteron, Panderichthys and Acanthostega), many of the fossils are literally fragments. The animals found represent only a small, stochastically selected, possibly quite unrepresentative, sample of the biodiversity that existed in these groups at those times. There is no way of knowing to what extent, if at all, those specific organisms were relevant to later developments, or what their relationships might have been to each other. They may all have been parts of lineages that died out, making no contributions to the overall flow of vertebrate evolution” (p. 341).
The geographic distribution of the available fossils does, however, favor multiple origins for tetrapods: “The geographic distribution of the relatively contemporaneous (within a few million years) known late Devonian tetrapods (east Greenland, eastern North America, northern Europe, Russia, and Australia), in the context of the distribution of the continents during the late Devonian, indicates high probability of genetically independent parallel origins of the different groups in multiple regions separated from each other by very long distances (varying from a few thousand km to, in the most extreme case, about 20,000 km). The chances that intervening environmental discontinuities and barriers were present seems very large” (p. 341).
Gordon concludes that “the applicability of the concept of monophyly at the macro-scales of evolutionary differentiation increasingly appears to be severely limited. An operational definition of the concept does not seem possible at the macro-level. Indeed, the phenomenon of a monophyletic origin of the universal tree of life probably did not occur” (p. 343). Gordon then urges that “conceptually and ideologically less constrained perspectives . . . may result in more nuanced and more biologically realistic sets of applications of the concept of monophyly” (p. 343).
In other words, if the history of life isn’t a single tree, then we should be free to consider that it might have been a forest. (For more on Gordon, see his personal web page at http://www.lifesci.ucla.edu/bio/html/gordon.htm).
“Towards a new evolutionary synthesis”. Robert L. Carroll, Trends in Evolution and Ecology, 2000, 15(1),:27-32.
Large-scale evolutionary phenomena cannot be understood solely on the basis of extrapolation from processes observed at the level of modern populations and species—though that is the central tenet of Neodarwinism. So says Robert Carroll in this stimulating article, in which he calls for an expanded concept of the evolutionary synthesis.
Robert Carroll is in the department of biology and is curator of vertebrate palaeontology, Redpath Museum, McGill University, Canada. Darwinism, he notes, “was based on the primarily uniformitarian concept” that the processes behind observable variations are sufficient to explain all diversification in the living world. Yet Carroll rejects this widely held thesis. The data from his own field of palaeontology count against it, he argues, noting that the observed “large-scale patterns and rates of evolution are not comparable with those hypothesised by Darwin on the basis of extrapolation from modern populations and species.”
Using arguments familiar to regular readers of Origins and Design, Carroll explains that what we find in the fossil record is the opposite of Darwinian gradualism. Instead of the Darwinian emphasis on smooth and continuous divergence, we find discontinuities. Populations and species “are clearly distinct in all taxonomic groups.”
What is missing are the many intermediate forms hypothesised by Darwin, and the continual divergence of major lineages into the morphospace between distinct adaptive types (p. 27).
Carroll zeros in on the Cambrian explosion, calling it the “most conspicuous event in metazoan evolution.”
The extreme speed of anatomical change and adaptive radiation during this brief time period requires explanations that go beyond those proposed for the evolution of species within the modern biota. . . . This explosive evolution of phyla with diverse body plans is certainly not explicable by extrapolation from the processes and rates of evolution observed in modern species, but requires a succession of unique events (p. 27-28).
Carroll proceeds to the genetics of developmental biology and is particularly interested in Hox genes in metazoans. These are homeobox genes that code for specific proteins that activate other genes, thereby providing a mechanism for cell regulation: “The number of Hox genes, arranged in a cluster along a chromosome, is broadly comparable to the degree of complexity of the organism, with one in sponges, four to five in cnidarians, six to ten in most of the higher metazoan phyla, and up to 39, arrayed in four Hox clusters on different chromosomes, in mammals” (p. 28).
This type of genetic control architecture, Carroll points out, was unknown to both Darwin and the founders of the Neodarwinian synthesis:
More broadly, the study of Hox genes has revealed a far different pattern of genetic control over structural features, throughout metazoan evolution, than those hypothesised by either mendelian or population genetics. Long-term evolution is not simply the result of selection of alternative alleles controlling specific traits, or the progressive accumulation of new mutations in an additive fashion, as proposed by quantitative genetics. It is now recognized that development involves a hierarchy of genetic control, including the precise timing and the position of expression of the Hox genes themselves, and the regulation of a cascade of downstream genes, together with interactions with the products of other Hox genes, commonly producing broadly pleiotropic results in many tissues and structures. Evolutionary changes over a wide range of magnitudes can occur by mutations at any point in this genetic complex (p. 29).
The last sentence above is not well supported. The potential for mutations to affect these genetic complexes is a matter for research, and limited results to date give grounds for great optimism for advances in evolutionary theory.
Carroll goes on to discuss palaeogeography, treating insights from plate tectonics regarding environmental pressures affecting organisms. Finally he considers the potential for integration: “The present generation of evolutionary biologists, whether trained as palaeontologists, molecular biologists, or population biologists, now has the opportunity to integrate this information into a new synthesis, to guide evolutionary research and teaching in the next century” (p. 30). At several places in this paper, it is clear that Carroll is frustrated with colleagues who appear to be locked into traditional Neodarwinism. He quotes favourably the summary of Schichting and Pigliucci: “The time is ripe for evolutionary theory to actively integrate development into its conceptual and experimental arsenal, lest we squander a second opportunity at a true synthesis” (p. 31). The clear implication is that Carroll considers the much-heralded Neodarwinian synthesis as a squandered opportunity.
The final point covered by Carroll is the role of natural selection. Despite numerous reports of apparently fast morphological change, Carroll points out that they are not linked to unidirectional transformations. Hence, they do not provide evidence of a mechanism that can explain large-scale evolutionary phenomena. “The relatively rare events involving the origin of major new taxa or significant morphological divergence at the species level require much greater than normal consistency of directional selection” (p. 31).
Carroll’s paper is important and timely, signalling that at the end of the 20th century we may well see a more open acknowledgement that the reigning Neodarwinian orthodoxy fails to conform to the evidence.
– David J. Tyler
Douglas Erwin, “The Origin of Bodyplans,” American Zoologist 39 (1999):617-629.
For several years, paleontologist Douglas Erwin of the National Museum of Natural History in Washington, D.C., has been quietly challenging a central tenet of Neodarwinism: that the biological present is the key to the evolutionary past. The idea is that processes observed today provide analogues to the processes by which life evolved in the past—or, as geneticist Theodore Dobzhansky put it, that a “sign of equality” obtains between “the mechanisms of macro- and micro-evolution” (Genetics and the Origin of Species, 1937, p. 12). Dobzhansky qualified that statement, however, by noting that the equality of micro- and macroevolution could be established only as a “working hypothesis.” Today, more than 60 years later, Erwin argues that the unknown processes that brought about the origin of the animal body plans (i.e., the major architectural features distinguishing different kinds of animals), whatever those processes may be, are ones for which we have no modern analogues. For “the simple, empirical fact is that the establishment of new body plans is not a frequent event” (p. 626). Thus Dobzhansky’s working hypothesis has been proved false.
For this working hypothesis depends on the principle that, as Erwin describes it, “the nature of evolutionary process is the same at all times and places.” But, the Cambrian Explosion of animal body plans refutes this principle: “There is every indication that the range of morphological innovation possible in the early Cambrian is simply not possible today” (p. 626). Thus micro- and macro-evolution are “effectively decoupled” (p. 625), and biologists must now seek to understand “the perplexing question of why such innovation (setting up the basic types of animals) is clustered in the history of life” (p. 618).
Kazuo Kawano, “How Far Can the Neodarwinism Be Extended? A Consideration from the History of Higher Taxa in Coleoptera,” Rivista di Biologia / Biology Forum 91 (1998): 31-56
Japanese biologist Kazuo Kawano joins a growing number of skeptical biologists and paleontologists who doubt the adequacy of Darwin’s argument from micro- to macro-evolution (see the companion article in this section on Douglas Erwin). Kawano focuses on the lone figure used in Darwin’s On the Origin of Species (see Figure 1), which Darwin used masterfully to explain his theory. Beginning at the lowest taxonomic level–varieties within a single species–Darwin argued that over the course of several generations, the members of a variety may diverge to the point where they are no longer able to interbreed, thus producing a new species. Darwin then extended the time scale of the diagram to include many thousands, and finally, millions of generations. By the same process of small-scale divergence, he argued, eventually new genera, families, and orders would arise, to the highest ranks of the taxonomic hierarchy. To use modern biological parlance, macroevolution is simply microevolution extended over much longer time scales. As a result, the steady extrapolation of ordinary variation and selection should explain the full range of biological form.
Figure 1. The famous lone figure from the Origin of Species (1859, p. 116). Darwin applies this diagram first at the level of “species of a genus,” (represented by A-L). He then increases the time scale (vertical dimension) severalfold, so that A-L become “distinct families, or even orders” (1859, p. 125). Here, the macroevolution of the higher taxonomic categories, such as phyla, begins at the level of the species. Click to see larger version.
As Kawano writes, “If the grand scheme set by Darwin’s diagram and strengthened by Neo-Darwinist theory is true, the evolution of multi-cellular animals should start from the evolution of lower taxonomic categories, moving up and spreading into higher categories, and the highest categories should emerge toward the latest stages of evolution” (p. 34).
Drawing on his studies of the beetle order Coleoptera, however, Kawano argues that this “grand scheme” is not true. He notes that variation is fundamentally limited, “confined within the common generic scheme” and “common ontogenetic program” defining the beetle type (p. 49). Varieties of beetles do not, and have not, given rise to new genera; instead, the basic type or body plan was established first, and later variations occurred within that type.
Kawano then extends this argument to the emergence of animals themselves:
The biggest incident in the evolution of multi-cellular animals, i.e., the emergence of phyla, took place in a relatively short period of time in the distant past. This was followed in turn by the evolution of classes, orders and families, and in the recent 70 million years at least no new family of Coleoptera evolved (p. 50).
He then directly challenges the Neodarwinian view:
The fact that phyla, classes, orders, and families had evolved in the remote past and never evolved again suggests that higher taxa, once evolved as a framework of variation, have stayed unchanged from the time of their emergence to today. The facts and logic indicate that the morphological evolution of multicellular animals has not been a spreading process but a process of diminishing dynamics where the magnitude of evolutionary effects on morphology decreased with time. Evolution is not a process of micro variations accumulating to macro effects but of macro effects preceding micro variations (p. 50).
Kawano sums up his argument in a figure (see Figure 2) differing dramatically from Darwin’s. Here the major types of organisms are shown to arise first, with their range of variation diminishing over time. “To understand how these frameworks [basic types] were established,” he writes, “is the greatest challenge to evolutionary biology” (p. 51).
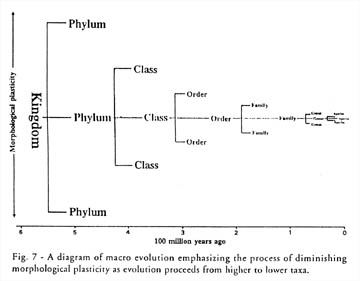
Figure 2.
Kazuo Kawano’s figure (1998, p. 52) showing “diminishing morphological plasticity”
over geological time. Contrary to the predictions of Neodarwinism, Kawano argues
that the basic body plans arise first, and any evolutionary elaborations occur
within those plans, with “genera” and “species” occuring to the far right. Thus,
variation is fundamentally limited by the boundaries of the basic plan or body
type. Click to see larger version.
Kawano concludes that Neodarwinism is not completely wrong, “but its applicability does not extend to evolution above the species level” (p. 52). The origin of the great patterns of life–those deep features distinguishing beetles from brachiopods–remains an open problem.
Francisco J. Ayala, “Molecular clock mirages,” BioEssays 21 (1999):71-75.
Is there a reliable molecular clock ticking away in the data of molecular biology--a measure by which evolutionary divergence can be estimated? No, says evolutionary biologist and geneticist Francisco Ayala (University of California at Irvine). In this review paper, Ayala surveys several recently published papers that “bring into question whether any predictive molecular clock hypothesis holds” (p. 72).
Many biomolecules deviate from the “stochastic clock” behavior (akin to radioactive decay) expected on the theory of neutral evolution from which the clock hypothesis derives, and to explain the divergences, various biological variables have been invoked, such as generation times, population sizes, or species characteristics. But Ayala argues that none of these attempted explanations is successful.
Looking at molecules such as superoxide dismutase (SOD), an enzyme that detoxifies free oxygen radicals, or glycerol-3-phosphate dehydrogenase (GPDH), an enzyme involved in animal metabolism, Ayala argues that none of the variables usually invoked can explain the strong divergence of SOD or GPDH from what the molecular clock predicts (see Table 1 for the rate divergences). “We are, then, left,” he writes, “with no predictive power and no clock proper” (p. 73). Ayala then examines studies of albumin and globin genes, as well as various genes from Drosophila sibling species, concluding that the “erratic evolutionary rates” (p. 73) and “erratic patterns of nucleotide evolution” (p. 74) cannot be explained by any molecular clock. “Without theoretical underpinnings to buttress the clock,” he concludes, “we will remain unable to anticipate when the clock might be erratic or how erratic it might be” (p. 75). While Ayala is not “willing to propose that we give up the clock altogether,” he counsels that “these and other inferences should be approached with caution” (p. 75).

Table 1.
Copyright 2000 Access Research Network. All rights reserved.
International copyright secured.
File Date: 6.27.01