January 1, 2005
The relaxing sound of a babbling brook; the happy laughter of a giggling child; the rousing sound of a marching band. All of these and more enrich our daily lives and are dependent on our ability to hear sound. But what exactly is sound and how is it that we are able to hear it? Keep reading to learn the answers to these questions and what logically follows as implications for the theory of macroevolution.
Sound! What’s it all about?
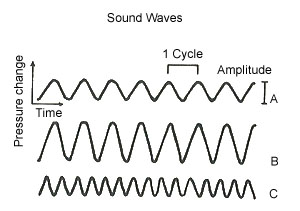
Sound is the sensation we experience when vibrating molecules of our surrounding environment (usually air), strike the ear drum. When these changes in air pressure, as determined by measuring the pressure on the tympanic membrane (ear drum), are plotted on a graph against time, a wave form appears. (see Figure 1). In general, the louder the sound, the more energy is required to produce it, and the greater the amplitude of air pressure change.
Figure 1. Wave B is louder than Wave A. Wave C is of a higher frequency than Wave A.
Loudness is defined by the decibel system using as its starting point the threshold for hearing (the level of intensity that something can just be barely heard by the human ear). The scale is logarithmic, which means that any jump from one absolute integer to the next, provided that it is divided by ten (don’t forget a decibel is only one-tenth of a bel) represents an increase in the order of ten times. For example, the threshold for hearing is designated as 0 and normal conversation occurs at about 50 decibels, so the difference in intensity is 10 raised to the power of 50 divided by 10 which equals 10 to the fifth power, or one hundred thousand times the intensity of threshold hearing. Or take for example a sound that causes you to feel severe pain in your ears and could be potentially damaging, which usually occurs at about the 140 decibel range; this sound, such as an explosion or a jet plane, would represent a 100 trillion-fold variation in sound intensity from threshold.
The shorter the distance between the waves, i.e. the more waves that are packed into one second of time, the higher the pitch, or frequency, of the sound being heard. This is usually designated as cycles per second (cps), or hertz (hz). (see Figure 1) The human ear is generally capable of hearing sounds that range in frequency from 20 hz to 20,000 hz. Normal human speech involves sounds from the frequency range of 120 hz for males, to about 250 hz for females. Middle C on the piano is 256 hz and tuning A done by the oboe for orchestras is 440 hz. The ear is most sensitive to sounds that range between
1,000- 3,000 hz.
A Concerto in Three Parts
The ear consists of three general regions designated as the external, the middle and the inner ear. Each plays its own unique and necessary part in allowing us to hear sounds.
Here’s a quick overview of which region does what and the components that are instrumental in accomplishing the function of hearing. (see Figure 2)

Figure 2. Anatomy of the Ear.
The pinna, or auricle, of the external ear acts like your own personal satellite dish by collecting and funneling sound waves to the external auditory meatus (opening to the ear canal). The sound waves then travel down the canal to the ear drum, or tympanic membrane, which by moving in and out in response to these changes in air pressure reproduces the vibration pattern of the sound source.
The three bones (ossicles) in the middle ear, called the malleus (hammer), which is directly connected to the tympanic membrane, the incus (anvil), and the stapes (stirrup), which is directly connected to the oval window of the cochlea, combine to transmit these vibrations to the inner ear. The middle ear is air-filled and is able to maintain the same air pressure on both sides of the tympanic membrane by way of the eustachian tube which connects up just behind the nose, and opens during swallowing to allow ambient air inside the middle ear chamber. Also, there are two skeletal muscles, the tensor tympani and the stapedius, which act to protect the ear from very loud sounds.
The inner ear, which contains the cochlea, first encounters these transmitted vibrations through the oval window, which results in a wave formation being set up in the internal structures of the cochlea. Within the cochlea sits the organ of Corti, which is the true organ of the ear, that is capable of converting these fluid vibrations into a nerve signal that is then sent off to the brain for interpretation.
So there you have it. Now let’s look at some specific aspects of each of these regions.
Say what?
Evidently, the external ear is where all of the action begins. If we didn’t have an opening in the skull that allowed sound waves to pass on to the ear drum, we wouldn’t be able to talk to each other. For some, maybe that would be considered a good thing! How exactly this opening in the bony skull, called the external auditory meatus, came into being by random genetic mutation or incidental change, remains to be explained.
The pinna, or ear flap if you will, has been shown to be important in sound localization. The underlying tissue that forms the pinna, allowing it to be so flexible, is called cartilage and is similar to the cartilage found in most of the joints of the body. How cells that are capable of cartilage formation acquired this ability, never mind how they ended up extending themselves from each side of the head, to the bane of many young women, would seem to require some sort of satisfactory explanation if one is to espouse the macroevolutionary model for the development of hearing.
Anyone who has ever undergone the experience of having had their ears plugged up with wax can appreciate the fact that although they may not know what benefit wax provides for the ear canal, they’re sure glad that it’s natural composition did not result in a substance that has the consistency of cement. Even more importantly, those who must interact with these unfortunate people appreciate their ability to elevate the volume of their voice in order to generate enough sound wave energy to be heard.
Ear wax, officially called cerumen, is a mixture of secretions from various glands contained in the external ear canal, which combine with the material from shedding cells along the lining of the canal, to form a white to yellow to brown waxy substance. Ear wax serves to lubricate the external ear canal while at the same time protecting the ear drum from dust, dirt, insects, bacteria, fungi, and anything else that the external environment can throw at it.
Interestingly enough, the ear has its own ear wax clearing mechanism. The cells that line the external ear canal form near the middle of the ear drum and migrate out to the walls of the canal and continue outward to the external auditory meatus. Along the way they carry with them the overlying ear wax which is then sloughed off when it reaches the outside opening. Jaw movements appear to enhance this process. In effect the whole scheme is like one big conveyer belt for wax elimination from the ear canal.
The whole understanding of wax formation, its consistency that allows for proper hearing, while at the same time providing an adequate protective function, and how the ear canal naturally eliminates it to prevent hearing loss, would seem to require some logical explanation. How could mere step by step innovations brought on by either genetic mutation or incidental change account for all of these factors and still allow for proper function along the way?
The ear drum, or tympanic membrane, consists of specialized tissue whose consistency, shape, attachments, and exact positioning, allow it be in the right place for the right function. It is all of these factors that need to be accounted for to explain how it is able to resonate in response to incoming sound waves and thereby start the chain reaction that results in the vibration wave within the cochlea. Just because other organisms have somewhat similar features that allow them to hear does not in itself explain how these features came into existence by the undirected forces of nature. In this I am reminded of the quip by G. K. Chesterton in which he said “It is absurd for the evolutionist to complain that it is unthinkable for an admittedly unthinkable God to make everything out of nothing, and then pretend that it is more thinkable that nothing should turn itself into everything.” But I digress.
Good Vibrations
The middle ear takes on the task of transmitting the vibrations of the ear drum to the inner ear where lies the cochlea in which is contained the organ of Corti, which is the actual “organ of the ear”, much like how the retina is the “organ of the eye”. So the middle ear is essentially the “middle man” in the operation of hearing. As often occurs in business, the middle man takes something away from the monetary efficiency of what is being transacted. So too, the transmission of ear drum vibration through the middle ear does result in some loss of energy resulting in only 60% of the energy being sent down the line. However, if it were not for the energy spread across the larger tympanic membrane being focused on the smaller oval window by the three ossicles, combined with their inherent lever action, this energy transmission would be much less and hearing would be much more difficult for us.
A projection from the malleus, (the first ossicle), called the manubrium, is directly attached to the ear drum. The malleus itself is connected to the second ossicle, the incus, which is itself attached to the stapes (the stirrup) which has a foot plate that is attached to the oval window of the cochlea. As mentioned already, the lever like actions of these three connected ossicles allow the vibration to become amplified on their way to the cochlea.
Review of two of my previous columns, namely “Hamlet Meets Modern Medical Science Parts I & II”, may allow the reader the see what is necessary to be demonstrated regarding bone formation itself. But how these three perfectly formed and interconnected bones ended up in precisely the right position to allow for the proper transmission of sound wave vibration, requires one more“ just-so” explanation of macroevolution to which we must look askance.
Curiously, within the middle ear exists two skeletal muscles, the tensor tympani and the stapedius. The insertion of the tensor tympani is attached to the manubrium of the malleus and on contraction it pulls the tympanic membrane back into the middle ear thereby limiting its ability to resonate. The insertion of the stapedius is attached to the the foot plate of the stapes and on contraction it pulls it off of the oval window, thereby reducing the amount of vibration that is transmitted to the cochlea.
Together these two muscles reflexively try to protect the ear from overly loud sounds which can cause pain and damage. The time that it takes for the neuromuscular system to react to a loud noise is about 150 milliseconds, which is about 1/6 th of a second. So sudden loud sounds, like from gunfire and explosions, are not as easily protected against as much as prolonged sounds or continuously noisy environments.
Experience tells us that loud sounds can sometimes be painful, just like overly bright light. The functional parts for hearing, such as the ear drum, the ossicles, and the organ of Corti, perform their function by moving in response to sound wave energy. Too much movement can cause damage and pain, just like if you overextend your elbow or your knee. So, it would seem that the ear has developed some sort of protection against self-injury if exposed to prolonged loud sounds.
Review of three of my prior columns, namely “Wired for Much More than Sound Parts I, II and III” which together explain neuromuscular function at a biomolecular and electrophysiological level, will allow the reader to better appreciate the inherent complexity that is contained within what would seem to be a natural protection against hearing loss. What is left to understand is how these two perfectly placed muscles ended up in the middle ear doing what they do, and doing them reflexively. What sort of genetic mutation or incidental changes occurred one step at a time to allow for such a complex development within the temporal bone of the skull?
Anyone, who on landing in an airplane, has experienced the sensation of pressure in their ears, associated with diminished hearing, and feeling like they are talking in a vacuum, has in effect demonstrated to themselves the importance of the Eustachian tube (auditory tube) that runs between the middle ear and the back of the nose.
The middle ear is an enclosed, air-filled chamber, in which the air pressure on either side of the tympanic membrane must be equal in order to allow for adequate mobility, which is referred to as its compliance. This is a measure of how easily the ear drum will move when stimulated by sound waves. The higher the compliance, the easier it is for the ear drum to resonate in response to sound, and the lower the compliance the more difficult it is to move in and out and therefore the threshold at which one can hear is raised i.e. now sounds have to be louder to be heard.
The air in the middle ear tends to be absorbed by the body which can result in the reduction of air pressure in the middle ear causing a reduction in tympanic membrane compliance. This occurs because instead of staying in the right position, the tympanic membrane will tend to be pushed into the middle ear by the ambient air pressure that is being exerted down the external ear canal since it is higher than the pressure in the middle ear.
The Eustachian tube connects the middle ear with the back of the nose and pharynx.
On swallowing, yawning and chewing, the associated muscular action tugs open the Eustachian tube which allows ambient air to enter and go up into the middle ear and replace any air that has been absorbed by the body. In this way, the tympanic membrane can maintain its optimal compliance which allows for adequate hearing.
Now, let’s go back to the airplane scenario. While you are cruising at 35,000 feet, the air pressure on both sides of the ear drum is the same, although the absolute amount is less than it would be at sea level. The important thing here is not the actual air pressure that is being exerted on either side, but that whatever the air pressure is on either side of the ear drum is the same. As you begin to descend, the ambient air pressure in the cabin begins to rise and immediately exerts itself against the ear drum from the external ear canal. The only way to correct this imbalance of air pressure across the ear drum is to be able to open up the Eustachian tube to allow the new ambient air pressure in. This is usually accomplished by chewing gum or sucking on candy that makes you swallow and apply that tugging action on the tube.
The speed at which the descent occurs and the resulting rapidly changing ambient pressure increases makes most people experience at least some sort of plugged sensation in their ears. If someone has or has recently had a cold, sinus problems or a sore throat, their Eustachian tube may not work as well during this pressure stressing event and they may experience severe pain, prolonged congestion and occasionally a severe hemorrhage in their middle ear!
But Eustachian tube dysfunction doesn’t end there. For if someone has chronic problems, over time the vacuum effect in the middle ear can pull fluid out of the capillaries which if not tended to can result in something known as glue ear. This is prevented and treated by myringotomy and tubes. The ENT surgeon puts a small hole in the ear drum and places tubes there so that any fluid that develops can migrate out of the ear and this serves to replace the Eustachian tube function until whatever has been causing it can be corrected, thereby preserving proper hearing and preventing damage to the structures within the middle ear.
It’s great that modern medicine is able to tackle some of these problems when the Eustachian tube doesn’t work right. But one has to immediately ask oneself how this tube came into being in the first place and which parts of the middle ear came first and how did it all function without the others? Does a step by step development based on some as yet unknown genetic mutation or incidental change even make sense here?
A close inspection of the parts of the middle ear and their absolute necessity for proper hearing that would allow for survival shows that there is an air of irreducible complexity about them. But none of what we’ve looked at so far will in itself result in us being able to hear. There’s one more piece of the puzzle to look at that has its own incredibly complex, and might I say, beautiful mechanism that takes the vibrations from the middle ear and converts them into a nerve message for the brain to interpret as sound.
Hardwired for Sound
The nerve cells that are responsible for sending the messages to the brain for hearing are located in the “organ of Corti” which is housed in the cochlea. The cochlea consists of three interconnected coiled tubes which spiral together for about two and a half turns.
(see Figure 3). The upper and lower tubes are both surrounded by bone and are called the scala vestibuli and the scala tympani respectively. Both of these tubes contain a fluid called perilymph whose sodium (Na+) ion and potassium (K+) ion contents are similar to other extracellular fluids (outside the cells) i.e. they have a high Na+ ion concentration and a low K+ ion concentration in contradistinction to intracellular fluid (inside the cells).
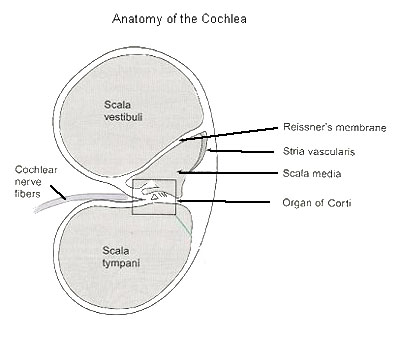
Figure 3. Anatomy of the Cochlea.
They communicate with each other at the tip of the cochlea through a small opening called the helicotrema.
The middle tube, which is embedded in membranous tissue, is called the scala media and it contains a fluid called endolymph which has the unique property of being the only extracellular fluid in the body that has a high concentration of K+ ions and a low concentration of Na+ ions. The scala media does not directly communicate with the other tubes and is separated from the scala vestibuli by flexible tissue called Reissner’s membrane and from the scala tympani by a flexible basilar membrane. (see Figure 4)
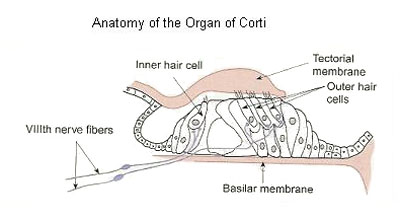
Figure 4. Anatomy of the Organ of Corti.
The organ of Corti sits suspended, like the Golden Gate Bridge, on the basilar membrane that is located between the scala tympani and the scala media. The nerve cells for hearing, called hair cells, because of their hair-like projections, sit on the basilar membrane which allows the bottom of the cells to be in contact with the perilymph of the scala tympani. (see Figure 4) The hair-like projections of the hair cells, which are known as stereocilia, sit on top of the hair cell and therefore are in contact with the scala media and the endolymph contained within it. The significance of this will become more apparent when we come to discuss the underlying electrophysiological mechanism behind auditory nerve stimulation.
The organ of Corti consists of about 20,000 of these hair cells that sit on the basilar membrane which runs for the entire spiraled cochlea, a distance of about 34 mm. In addition, the thickness of the basilar membrane varies from about 0.1mm at the beginning, the base, to about 0.5mm at the end, the apex, of the cochlea. This feature will become important when we discuss pitch or frequency.
Now remember, sound waves have entered the external ear canal where they have caused the ear drum to resonate at an amplitude and frequency that is inherent within the sound itself. The inward and outward motion of the ear drum allows vibration energy to be transferred to the malleus, which is connected to the incus, which is in turn connected to the stapes. In the ideal circumstance, the air pressure on either side of the ear drum is equal, allowing for the ear drum to have a high compliance for motion, because of the Eustachian tube’s ability to allow ambient air into the middle ear from the back of the nose and throat when yawning, chewing and swallowing occur. This vibration is now transferred from the stapes to the cochlea via the oval window. Now we’re ready for action.
The resulting transfer of vibration energy to the cochlea causes a fluid wave to be transmitted through the perilymph in the scala vestibuli. However, because the scala vestibuli is encased in bone and is separated from the scala media, not by a rigid wall, but by a flexible membrane, this vibration wave is also transmitted to the endolymph in the scala media by way of Reissner’s membrane. The resulting fluid wave in the scala media is itself responsible for causing the flexible basilar membrane to also undulate. These waves peak and then die down quickly somewhere along the basilar membrane in direct relationship to the frequency of the sound being heard. The higher frequency sounds cause more motion at the base or thinner part of the basilar membrane, and the lower frequency sounds cause more motion at the apex or thicker part of the basilar membrane, at the helicotrema. Eventually the wave action comes into the scala tympani via the helicotrema and dissipates through the round window.
One can immediately see that if the basilar membrane is waving in the “breeze” of endolymphatic motion within the scala media, that the suspended organ of Corti, with its hair cells, is going to undergo a trampoline-like effect in response to this wave motion energy. From here on, in order to appreciate the complexity and to truly understand what is going on for hearing to take place, the reader must have a knowledge of neuron function. If you haven’t read it already, I suggest that you look at “Wired for Much More than Sound Parts I and II” which reviews neuron function.
Hair cells at rest have a membrane potential of about -60mV. Remember from neuron physiology that the resting membrane potential exists because of the tendency for more K+ ions to leave the cell through K+ ion channels than Na+ ions entering through Na+ ion channels when the cell is not stimulated. However, this tendency is predicated on the fact that the cell membrane is in contact with extracellular fluid that is usually low in K+ ions and high in Na+ ions, like the perilymph with which the base of the hair cells are in contact.
When the stereocilia, i.e. the hair-like projections of the hair cell, are stimulated to move by wave action, this causes them to bend. This motion of the stereocilia results in certain transduction channels being opened that are very permeable to K+ ions. So when the organ of Corti experiences this trampoline-like effect from the wave action brought on by the vibration from the resonance of the ear drum through the three ossicles, this results in K+ ions entering the hair cell, which causes it to depolarize, i.e. become less negative in its membrane potential.
“Hold it”, I hear you say. “I just reviewed all of that stuff on neurons and to my way of thinking when the transduction channels open up, K+ ions should flow out of the cell and cause hyperpolarization, not depolarization.” And normally you’d be absolutely right because in the usual set of circumstances when specific ion channels open up to increase the permeability of that specific ion across the membrane, it is Na+ ions that go into the cell and K+ ions that come out because of the relative concentration gradients of Na+ ions and K+ ions across the membrane.
But remember, we’re not dealing with the usual set of circumstances here. The apex of the hair cell is in contact with the endolymph of the scala media and not the perilymph of the scala tympani that is contact with the base of the hair cell. And remember, we stressed the point above that the endolymph has the unique distinction of being the only fluid outside of the cell that has a high concentration of K+ ions. So high that when those transduction channels that are permeable to K+ ions open up in response to the bending motion of the stereocilia, K+ ions now enter the cell and thereby causes it to depolarize.
The depolarization of the hair cell causes voltage-gated calcium ion (Ca++) channels in its base to open up and to allow Ca++ ions into the cell. This results in a neurotransmitter from the hair cell being released to stimulate a nearby cochlear neuron which will ultimately send the message on to the brain.
The frequency of the sound that generates the fluid wave determines where it will peak along the basilar membrane. As mentioned above, this is dependent on the basilar membrane’s thickness, in which higher pitched sounds cause more activity at the thinner base, and lower frequency sounds result in more activity at the thicker apex.
One can immediately see that the hair cells that are closer to the base will maximally respond to very high pitched sounds at the upper limit of human hearing (20,000 hz) and the hair cells at the extreme apex will maximally respond to sounds at the lower limit of human hearing (20 hz).
The cochlear nerve fibers demonstrate tonotopic mapping in that they are more sensitive to specific frequencies which are ultimately mapped out in the brain. This means that specific cochlear neurons service specific hair cells, and their nerve signals are eventually transmitted to the brain which is then capable of determining the pitch of the sound based on which hair cells were stimulated. In addition, it has been shown that cochlear nerve fibers have spontaneous activity so that when they are stimulated by a sound of a specific pitch with a particular amplitude, this results in a modulation of their activity which is ultimately analyzed by the brain and is interpreted as a particular sound.
In summary, the hair cells that are located on a specific spot on the basilar membrane will maximally bend in response to a particular pitch of sound wave that results in that spot on the basilar membrane receiving the crest of the wave. The resulting depolarization of that hair cell will cause it to release a neurotransmitter which will stimulate a nearby cochlear neuron that sends its message along to the brain where it is interpreted as the sound that was heard with a certain amplitude and frequency based on which cochlear neuron sent the message.
The pathways for all of this auditory nervous activity have largely been mapped out. There are more neurons that are contained in junction boxes that receive these messages and then pass them on to other neurons. Eventually the messages reach the auditory cortex of the brain for final analyses. But how the brain then converts this myriad of neurochemical messages into what we know as hearing is as yet totally unknown.
The impediments to solving this problem may indeed be as mysterious as life itself!
Sound Advice?
A brief review of cochlear structure and function will provide the reader with many questions to be asked of those who are enamored with the theory that all life came about through the random forces of nature without any intelligent input. Here are a few of the major factors whose development over time are in need of plausible explanations given their absolute necessity for the function of hearing in humans.
Is it possible that these developed one step at a time by the processes of genetic mutation or incidental change? Or failing that, is it likely that each of these parts served some, as yet unknown, function in multiple other progenitors, which then came together to allow for human hearing as we know it?
And if either of these explanations are considered valid, then what exactly were these changes in principle, and in fact, to allow for the development of such a complex system that allows for the transduction of air waves into something that the human brain perceives as sound?
The development of the three coiled tubes, called the scala vestibuli, the scala media and the scala tympani, which form the cochlea
The presence of the oval window to receive the vibration from the stapes and the round window to allow the wave action to dissipate
The presence of Reissner’s membrane to allow the vibration wave to be transmitted to the scala media
The basilar membrane, with its variable thickness and its perfect location between the scala media and the scala tympani, to play a part in the function of human hearing
The construction of the organ of Corti and its position on the basilar membrane so that it may experience the trampoline effect that is instrumental for human hearing
The presence of the hair cells within the organ of Corti whose stereocilia play an all important role for human hearing and without which, it would not exist
The presence of perilymph in the upper and lower scalae and endolymph in the scala media
The presence of the cochlear nerve fiber in close proximity to the hair cells that are located on the organ of Corti
A Final Word
When I set out to write this column I first looked at the original medical physiology text that I used when I was in medical school 30 years ago. In this book the authors noted the unique make-up of endolymph as compared to all of the other extracellular fluids in the body. At this point in time, it was “unsettled” as to the exact reason for this unusual set of circumstances and the authors freely admitted that although it was known that the action potential that was generated by the auditory nerve was related to the movement of the hair cells, how this happened was as yet unknown. So what are we to make of this now that we have a better understanding of how all of this works? Simply this:
Is there anyone who upon listening to their favorite piece of music thinks that the notes being played in that specific order have come about by the random forces of nature?
No! One realizes that the music one is enjoying was written down by a composer so that others would be able to enjoy what he had created and heard in his own mind. He makes sure of this by signing his own name to the original manuscript so that the world will know who created this piece. To think otherwise would result in most people being subjected to ridicule.
Similarly, as one listens to the cadenza being played in a violin concerto, is it likely to come into one’s mind that the notes emanating from that Stradivarius have occurred simply by the random forces of nature? No! One intuitively knows that there is a talented virtuoso who is playing specific notes in order to produce the sounds that she wants the listener to hear and appreciate. So much so, that her name is emblazoned on the CD jacket so that those who know of her talent will be induced to buy it.
But how is it that one is able to hear what is being played in the first place? Did it all come about by the undirected forces of nature as evolutionary biologists believe? Or is it possible that at some point in time an intelligent designer made its presence known, and if so, how would we be able to detect it? Are there any signatures or emblazoned names within nature that may help direct our attention to them?
There are numerous examples of what I consider intelligent design within the human body which I have detailed over the last year within this column. But when I came to the realization of how hair cell motion results in the opening of K+ ion transduction channels causing the movement of K+ ions into the hair cell and its depolarization, I was literally dumbfounded. For I suddenly realized that here was such a “signature”. Here was an example of the intelligent designer letting it be known that just when humanity thinks it knows all there is to know about life and how it developed, it is faced with something that should give it pause.
Remember that the almost universal mechanism for neuron depolarization occurs by the influx of Na+ ions from extracellular fluid into the neuron through Na+ ion channels after sufficient stimulation. The development of this system in itself has as yet not been sufficiently explained by evolutionary biologists. However, the whole system depends upon the existence and stimulation of Na+ ion channels in combination with there being a higher concentration of Na+ ions outside of the cell as compared to inside the cell. This is how the neurons of the body work.
Now we come to find out that there exists within the body a set of neurons that work in exactly the opposite way. They require, not Na+ ions to enter the cell to cause depolarization, but K+ ions. On the face of this, it would seem to be an impossibility because everyone knows that all extracellular fluids in the body contain very low amounts of K+ ions in comparison to the inside of the neuron and therefore it would be physiologically impossible for K+ ions to flood into the neuron to cause depolarization the way that Na+ ions do.
What was once considered as “unsettled” has now become crystal clear as to the reason why endolymph must have the unique property of being the only extracellular fluid in the body with a high K+ ion content and a low Na+ content. It is located precisely in the right place so that when the K+ ion permeable transduction channels open in the membrane of the hair cells, depolarization will take place. Evolutionary biologists must be able to explain how both, seemingly opposing sets of circumstances, could arise and how they could occur in the right place in the body where it is needed. Much like the notes for a concerto being placed just right by the composer and then being played on the violin by the virtuoso. To me, that’s an intelligent designer saying to us; “Do you see the beauty in what I have created?”
Of course, for a person who sees life and how it functions only through a materialistic and naturalistic filter, the idea of an intelligent designer is an impossibility. The fact that all of the questions that I have proposed for macroevolution in this and other columns, are highly unlikely to receive plausible answers in the future, does not seem to deter or even concern the proponents of the theory that all life has developed from natural selection acting on random variation.
As William Dembski so adroitly observed in The Design Revolution: “Darwinists take this present lack of insight into the workings of an unevolved designer, not as remediable ignorance and not as evidence that the designer’s capacities far outstrip ours, but as proof that there is no unevolved designer, period.”
Next month we’ll be looking at how the body coordinates its muscular activity in order to allow us to sit, stand, and stay mobile: in my last installment on neuromuscular function.
See you then in: Wired for Much More than Sound Part VIII: Run for your Life
Dr. G.
Howard Glicksman M. D. graduated from the University of Toronto in 1978. He practiced primary care medicine for almost 25 yrs in Oakville, Ontario and Spring Hill, Florida. He recently left his private practice and has started to practice palliative medicine for a Hospice organization in his community. He has a special interest in how the ethos of our culture has been influenced by modern science’s understanding and promotion of what it means to be a human being.
Copyright 2004 Dr. Howard Glicksman. All rights reserved. International
copyright secured.
File Date: 1.01.05