November 1, 2004
Last month’s column dealt with the many parts of the eye and how they, together, are able to allow light to enter the eye and be focused on the retina. This month we look at how the retina is able to accomplish its task of converting light into a message that is eventually interpreted by the brain as vision. Here’s an overview of what we’ll be talking about in this column.
The retina is a multi-layered sheet of neural tissue that is sensitive to light which makes up the inner lining of the back part of the eyeball. The retina sits on a pigmented membrane, known as the retinal pigmented epithelium (RPE), which absorbs any light that passes through the retinal tissue. This prevents the light from reflecting back through the retina and causing the retinal cells to react again, thereby preventing visual blurring.
Light travels through the eye and creates a complicated chemical reaction in the light-sensitive photoreceptor cells of the retina, (rods and cones) which results in a change of neuron function. The RPE also plays a major role in the recycling and regeneration of the materials that are necessary for photoreceptor function which are contained in the rods and cones.
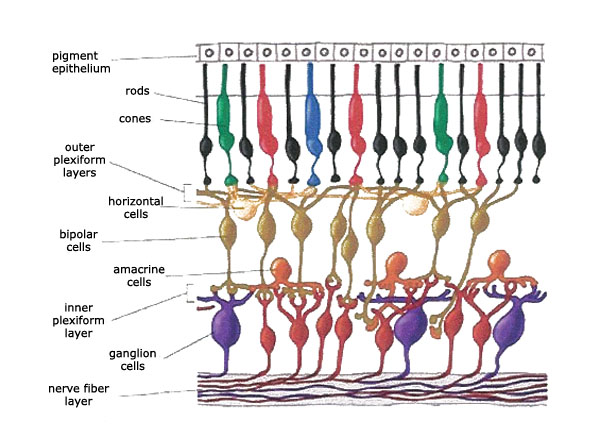
The alteration of rod and cone cell function brought on by being exposed to a photon of light is only the beginning of the nervous impulse that eventually reaches the brain as vision. The rods and cones are not in direct contact with the visual cortex of the brain. They send their messages to bipolar cells which then send their messages to the ganglion cells. It’s the axons of the ganglion cells that make up the optic nerve that winds its way to the brain.
But that’s not the full story either. For there exists two layers of interneurons. The horizontal cells relay messages back and forth between the photoreceptor cells, the bipolar cells and each other. And the amacrine cells are interconnected with the bipolar cells, the ganglion cells and each other as well. Both of these types of interneurons play a major role in the processing of visual information at the level of the retina before it is passed on to the brain for final interpretation. (See Figure1)
Figure 1. Retinal Cells. Obtained by the permission of the Scientific Research Society.
We Live for Light
There are about 120 million rod cells in the retina of the eye. These photoreceptor cells contain a photopigment called rhodopsin, consisting of two separate molecules called retinal, and a membrane based group called, an opsin. Rhodopsin is dependent on Vitamin A and is very sensitive to all wavelengths of the visible light spectrum.
The rods are strewn throughout the retina except that there are very few in the macula, and none in the fovea. The rods provide us with our night and peripheral vision and allow us to discern between light and dark objects.
In contrast to the rods, there are only about 6 million cones in the retina and they are mainly concentrated in the macula, with the highest amount in the cone-only fovea. They require brighter light than the rods to function properly and they provide us with our color and central vision which allows us to read and focus on things.
In humans, there are three different types of cone cells, each of which contains a photopigment that is rhodopsin-like in nature and structure. This is each chemically connected to an opsin that is a little different from the one in rhodopsin. These changes in the opsin molecule of the cone cells results in each of them being more sensitive to a particular wavelength of light. Although all of the cones will react somewhat to the entire spectrum of visible light, each cone will have a predilection to react more strongly to one specific wave length. This is how we are able to see the colors, red, green and blue and the combinations thereof.
Recently I happened across an article called “Where Would an Eye Spot Have Come From?” in which the simplicity of the development of photosensitive receptors was assumed only because it was known that light sensitive chemistry existed in more primitive organisms. Actually the photosynthesis of plants is accomplished by chlorophyll which is totally different in structure from rhodopsin. However there are bacteria that produce a rhodopsin-like molecule that enables the bacteria to absorb the energy from light and use it for functioning.
What the author neglected to tell us is how this bacterial rhodopsin could have come into being in the first place. First one needs to explain the development of the two separate molecules that are necessary for its formation and one also needs to take into account that the opsin molecule consists of over 300 amino acids arranged in a specific order. Since there are 20 amino acids to choose from, the chances of forming a protein molecule with 300 amino acids in the right order to allow for photosensitivity is one chance in 10 raised to almost the 400th power. Then you have to explain how the cell has the underlying information (blueprint) and machinery to create this specific molecule for a specific purpose of function.
The mere presence of rhodopsin-like molecules in other organisms does not prove that they came about by the random forces of nature. It only proves that there are similar biomolecular structures that function in similar ways within various organisms. And why should we be so surprised since all of life is under the same constraints of the physical and chemical laws of the universe? It’s like assuming that just because a lawn mower and an automobile both need oil and gas and use them in a somewhat similar fashion, that they must have come into being by the random forces of nature. Are we to suppose that as long as there are similarities between the biomolecular structures and functions of different organisms that this must only mean that they developed from each other by the random forces of nature? But all of human invention empirically tells us that this may not be so. And therefore the world must demand to know the specifics of the mechanism behind these incredibly complex transformations attested to by macro-evolution and its supporters.
In addition, the author stated “We can assume that something like Vitamin A was around, since it is found in so many creatures and rhodopsin is just a variation of Vitamin A.” Now although the bacteria that produces its own rhodopsin may indeed be able to produce Vitamin A, the very fact that it is a vitamin tells us that humans cannot. This would seem to make humans more inefficient in that they would have to be dependent on obtaining their own Vitamin A from outside sources. If Vitamin A production already existed in the armamentarium of biological life, why didn’t we develop the capacity to produce our own if it is so important for vision? How did we develop the ability to produce rhodopsin but not the ability to produce the Vitamin A that is absolutely necessary for it in the first place?
So, although rhodopsin may have existed in more primitive organisms like the bacteria, that really doesn’t help humans that much because unless we can get a hold of some Vitamin A we’re not going to be able to produce our own rhodopsin. The absorption of Vitamin A by the gastrointestinal system and its subsequent metabolism in the body is a very complex process that involves yet more biomolecules and organ structures, and requires it own explanation of development by macroevolution. There are many other functions of the body besides vision that are vitally dependent on Vitamin A, namely, skin and mucous membrane development and growth, and immune function.
Although some bacteria may be able to produce their own Vitamin A for their rhodopsin, the same cannot be said of humans and therefore any discussion that purports to explain the development of Vitamin A dependent rhodopsin production in humans that does not at the same time detail the development of the body’s ability to absorb and metabolize Vitamin A in a manner that allows for human survival should be considered deficient and dare I say…..well….when properly understood, actually unscientific!
These particulars are never mentioned in this article, so that the reader may not notice that a rational and physiologically sound explanation of how these biomolecules could have developed one step at a time and could have been maintained in a physiologically sound manner, is actually absent from the discussion. As more and more becomes known about how the body functions and how easily it can breakdown, it becomes more and more evident that almost all of the vital functions of the body are indeed vitally inter-dependent. One can’t suppose to have explained how one particular organ came into existence unless one has also explained the development of all of the rest of them in what would seem to have had to have been a simultaneous event. Maybe this occurred during the biological big bang that is spoken of, that occurred about 500 million years ago when suddenly, about fifty new multi-system organisms with complex body plans came into existence?
Retinal Relations
As mentioned already, once the photoreceptor cells react to a photon of light, they don’t send their messages directly to the brain. In fact, the message then gets sent to a bipolar cell and then onto a ganglion cell which then sends it to the brain. The horizontal cells and the amacrine cells are able to modify the final message that is sent along the optic nerve and in effect they greatly contribute to the retina’s ability to process these messages and at least, in part, construct the visual image that will ultimately be detected in the brain.
Central vision is dependent on the cone-dense fovea and the light that is focused on it by the combination of the cornea and the lens, with or without accommodation by the ciliary muscle. The neural message from each cone in the fovea is attended to by a single bipolar cell which itself is serviced by one ganglion cell. In this way, there is a 1:1:1 relationship between the cones of the fovea and the bipolar and ganglion cells that receive their messages. Therefore, the message from each cone in the fovea is not combined with any other cells in the retina and results in the brain receiving highly specific neuro-excitatory information from each cone cell, which gives us sharp vision.
The rod cells however do not have the luxury of having their own private bipolar and ganglion cells to receive their messages. In fact, in the outer reaches of the retina, one bipolar cell might service over 100 rod cells. And then the ganglion cell responsible for picking up the messages from the bipolar cells out in the boondocks of the retina may be receiving messages from more than one bipolar cell. Also, rod cells sometimes share their changes in neural function by way of gap junctions and sometimes a rod cell is capable of stimulating several different ganglion cells.
This situation is borne out by the fact that although there are about 120 million rod cells and 6 million cone cells in each eye, there are only about 1.2 million ganglion cell axons that make up the optic nerve. So it is evident that each photoreceptor cell cannot have a direct impact on every ganglion cell like the cone cells in the fovea.
Taking into consideration how rod cell transmission eventually reaches the brain, it is evident that when a given rod cell stimulates a bipolar cell that services more than a 100 others, and then passes its message on to a ganglion cell which in turn receives messages from other bipolar cells, that the brain will have no way of knowing exactly which rod cell fired although it will be able to let you know which region was involved. This is why our peripheral and rod cell vision is not as refined and specific as our central vision, in fact it’s downright blurry and out of focus.
Focusing is Not Enough
Last month’s column critically looked at the claim by macroevolution that the human eye, with all of its interdependent and biologically complex parts, could have developed one step at a time by the random forces of nature. Included in that exercise was a detailed description of how the eye is able to focus incoming parallel and divergent light rays onto the fovea for us to have central vision. A review and analysis of what you’ve just read should make you realize that the focusing of light on the fovea is not in itself adequate to afford us the ability to see things that interest us so clearly. It is only the first
of three necessary elements that have been described here and in last month’s column.
Central vision is clearly dependent on not only the eye’s ability to focus light on the fovea, but also that the fovea contain an adequate supply and concentration of appropriate photoreceptor cells that will react to the focused light in a way that has a chance of resulting in central vision. But even that is not enough to allow us to be able to focus in on objects of interest.
What would happen to our central vision if the cone cells of the fovea were connected to the bipolar cells and then the ganglion cells the way the rods are? If many cones sent their vital messages to only one bipolar cell and it in turn joined other bipolar cells to stimulate a ganglion cell which would eventually send a message along to the brain, it is evident that the brain would have the same problem in precisely identifying which part of the fovea had been stimulated and the image would be distorted and blurry.
No! Our ability to be able to clearly see objects of interest that allow us to read, create, interact, observe, avoid, and ultimately imagine, is due not only to the cornea and the lens in tandem being able to focus light on the fovea, but also the high concentration of cone cells in the fovea and the 1:1:1 cone:bipolar:ganglion cell connection which allows the brain to receive enough highly specific information about what we’re observing to allow us to see it clearly.
One must wonder which of these innovations came on the scene first? Last month’s column highlighted the fact that many of the assumptions of how the cornea could have developed over time by the random forces of nature do not take into account the underlying physiology that keeps the cornea alive and functioning i.e. its dependence on the secretions of the lacrimal gland, the glands in the conjunctiva, and the ciliary body’s production of aqueous fluid. But how would an organism be benefited by having the ability to focus a cone of light on a particular region of the retina if there weren’t the appropriate amounts or types of photoreceptors there to receive the message in the first place? And what would be the use of having a cone-dense fovea if the eye were incapable of focusing the light on it?
But even if the eye, at some stage along the way of its development, were capable of directing the light from an object of interest into something like the fovea, what use would it be if the neurocircuitry servicing the photoreceptor cells did not provide for enough specific information to be sent to the processor in the brain? And of course, the opposite could be asked as well; if the 1:1:1 neuronal connections to the brain were in place, what good would it do the organism if its eye couldn’t focus light properly or it didn’t have a cone-dense fovea? So, which came first and how did it benefit a multi-system organism with a complex body plan without the other two? Just something to think about!
The Pause that Refreshes
Before going any further I would advise anyone reading this who is not familiar with neurons and how they function to review my two prior columns that deal with this issue. (Wired for Much More than Sound Parts I and II, Neurons and how they Work, The Impulse and The Transfer). Without this knowledge you are likely to have great difficulty in following what is discussed going forward, and even more importantly, you are liable to lose some appreciation for the complexity and sophistication of neuro-ophthalmological function in humans.
For without a firm understanding of what is occurring at the biomolecular and electro-physiological levels, you could easily be misled into believing that it would be reasonable to assume, that after the step by step development, by natural processes, of a precursor primitive eye spot, as macroevolution proposes, that nervous and muscular connections arose allowing for a brain-like structure to interpret its messages as vision while at the same time controlling its movement.
These descriptions that purport to explain how all of this happened; “Which came first the eye or the brain?”; are replete with optimistic and simplistic explanations, which are surrounded by rhetorical language, but are devoid of any detailed scientific description of how the myriad of biomolecular reactions involved and the associated genetic material necessary for their substrate, came into being, and how the neurocircuitry and its underlying electrophysiology developed over time. Certainly these explanations may serve to satisfy the intellects of those who are committed to the belief of macroevolution, but to me they seem to border on what is commonly called science fiction.
But How Does it Really Work?
More and more is being learned about the retina, but as yet there are still many questions that need to be answered. It is known however that when a photon of light, which is a packet of electromagnetic energy, interacts with rhodopsin in the rod cell, that this results in it changing its shape. The net effect of this transformation is a series of very rapid chemical changes taking place in the opsin component of the molecule which alters its chemical properties.
This change in rhodopsin starts a chemical reaction within the photoreceptor cell involving three biomolecules; namely, transducin: which activates the enzyme phosphodiesterase (PDE): which breaks down cGMP (3’5’ guanosine mono-phosphate). The drop in cellular cGMP results in the closing of the Na+ and Ca++ ion channels along the cell membrane which hyperpolarizes the photoreceptor’s membrane potential. You may recall that when a neuron’s membrane potential becomes more negative, this causes inhibition, and for the photoreceptor cell this means the slowing down or complete stoppage of the release of its neurotransmitter, glutamate.
Note that this hyperpolarizing response to light that results in the decrease of neurotransmitter release occurs only in vertebrate eyes. Most invertebrate photoreceptor cells depolarize, or are excited, when exposed to light, which will result in an increase of neurotransmitter being released, much like the motor neuron. If you’re scratching your head wondering how this hyperpolarizing action would allow us to see, get ready to wonder as you’ve never wondered before.
It appears that the rods and cones do not actually produce action potential spikes like other nerve tissue. In the dark, they are continuously releasing glutamate which in this setting is inhibitory. This means that in general, the rods and cones usually are sending inhibiting messages to their interconnecting neurons. So when a photon of light strikes the photoreceptor and closes its Na+ and Ca++ ion channels result-ing in hyperpolarization of the cell membrane, this causes the cell to decrease its output of an inhibitory neurotransmitter (glutamate). Essentially this is like a double negative that results in a positive response. The interconnecting cell which has been previously inhibited by the secretion of glutamate by the presynaptic photoreceptor cell has been released from that inhibition and is free to start sending its own message down the line.
In general, the photoreceptors, the horizontal cells, the bipolar cells and most of the amacrine cells do not have action potentials that result in an “all or nothing” response. They have the ability to release their neurotransmitters at certain voltage levels which can be modulated by the impact of the incoming neurotransmitters released by each other. In other words, the release of glutamate will often cause hyperpolarization of a postsynaptic cell and diminish its release of neurotransmitter.
There are many different types of neurotransmitters that are released by different types of interneurons which can either inhibit (hyperpolarize) or excite (depolarize) the postsynaptic cell. The horizontal cells are able to communicate with each other and also sends messages back to the photoreceptors and on to the bipolar cells. Similarly, the amacrine cells seem to be able to contact and affect the actions of other amacrine cells and both bipolar and ganglion cells. These interactions involve many different types of neurotransmitters and the nature of these modifications and their significance for the processing of the visual message before it is sent to the brain is at present under study.
Of note is the fact that although, generally, glutamate is thought to be an inhibitory neurotransmitter that results in the hyperpolarization of the postsynaptic cell which causes a diminution of that cell’s neurotransmitter release, it has been now shown that some bipolar cells react in the opposite way because they have a different gluatamate receptor protein on their membrane and are actually depolarized (excited) by their interaction with glutamate.
It has been determined that there exists an ON pathway and an OFF pathway in the retina. All bipolar and ganglion cell input is generated from a receptive field pattern that comes under one of these two designations. Keep in mind that all photoreceptor cells continue to release the neurotransmitter, glutamate, when totally in the dark.
The ON pathway involves bipolar cells that are turned “on” when the photoreceptor cells are exposed to light. Therefore in the dark, while the rods and cones are releasing maximal amounts of glutamate, these bipolar and ganglion cells are inhibited. This in effect keeps the ON pathway turned off. But when the light comes on and interacts with the photopigment in the photoreceptor cell, this reduces the release of glutamate from the rods and cones. The suppressive effect on the bipolar and ganglion cells is removed and they are able to send their messages along. Therefore, when the light turns on, the ON pathway turns on as well. This system allows us to be able to see light objects on dark backgrounds.
The OFF pathway works in the opposite way. When the photoreceptor cells are in the dark and are releasing maximal amounts of glutamate, this depolarizes (excites) the OFF bipolar cells and ganglion cells. This in effect allows the OFF pathway to stay on when the rods and cones are in the dark. But when light strikes the photoreceptors servicing the OFF pathway, the reduction in glutamate release reduces the output from the OFF bipolar and ganglion cells resulting in them turning off. So when the light is on, this system turns off. It is the OFF pathway that allows us to see dark objects against light backgrounds. It’s most likely the OFF pathway that you are using right now as you read this column. (See Figure 2)

Figure 2. The ON and OFF pathways . Obtained by the permission of the Scientific Research Society.
There are many other levels of complexity that exist within the receptive fields of the retinal neurocircuitry that have been determined to exist and have subsequently brought on other questions for scientists. For example, within a given ON and OFF receptive field it is known that if light hits the center of the field that it will turn the bipolar and ganglion cells ON or OFF respectively. But if it hits the circular region known as the surround (think of it like a target with a bull’s eye) then this region will turn OFF and ON respectively. In other words, the surround region reacts in the opposite way of the center region for which the particular ON or OFF pathway is named. This allows us to be able to contrast between light and dark objects. i.e. see light objects on a dark background and dark objects on a light background. (See Figure 3)

Figure 3. The center and surround receptor fields. Obtained by the permission of the Scientific Research Society.
But Still More is to be Revealed
All of this and more makes the retina a treasure trove for anyone who is fascinated by neurobiological function and is interested in possibly providing humankind with more information about how it works. Isn’t it interesting that modern science still doesn’t know all that there is to know about how the retina works, but it seems pretty certain about how it came into existence, one step at a time, by the random forces of nature? One could say that knowing that all of the evidence has yet to be discovered about retinal function would seem to make any judgment about the origin of the eye, its connections to the brain, and ability to experience vision, appear to be a bit premature, even hasty. The jury is still out on origin theories, despite what modern science may say.
Does macroevolution make sense given this information, or does intelligent design, or does neither? Maybe, armed with this information you will be better able to critically analyze some of the claims of macroevolution and be able to ask some penetrating questions that require definitive answers.
Next month we’ll finish this three part series about vision by examining what happens to the optic nerve messages that are sent to the brain. What kind of neuroexcitatory pattern does the visual cortex receive and how does it turn it into what we call vision? Look forward to seeing you then with:
Wired for Much More than Sound- PartVI—Vision: Part 3—What Does the Brain See?
Dr.G
Howard Glicksman M. D. graduated from the University of Toronto in 1978. He practiced primary care medicine for almost 25 yrs in Oakville, Ontario and Spring Hill, Florida. He recently left his private practice and has started to practice palliative medicine for a Hospice organization in his community. He has a special interest in how the ethos of our culture has been influenced by modern science’s understanding and promotion of what it means to be a human being.
Copyright 2004 Dr. Howard Glicksman. All rights reserved. International
copyright secured.
File Date:11.01.04